

Research Article - (2022) Volume 6, Issue 1
Evaluation of Sinapic Acid to Ameliorate Ionizing Radiation Induced Peripheral Blood Mononuclear Cell Death
22Department of Physiology, Sister Nibedita Govt. General Degree College, for Girls, Hasting’s House., 20B Judges Court Road, Alipur, Kolkata 700027, west Bengal, India
Received Date: Apr 23, 2022 / Accepted Date: May 05, 2022 / Published Date: May 12, 2022
Copyright: ©Copyright: Ã?©2022 Sarmishtha Chanda, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited
Citation: Prasenjit Mondal, Adity Bose, Sarmishtha Chanda, (2022). Evaluation of Sinapic Acid to Ameliorate Ionizing Radiation Induced Peripheral Blood Mononuclear Cell Death. Stem Cell Res Int 6(1), 34- 45.
Abstract
Introduction: The UV radiation is known as a potential environmental carcinogen which causes damages of macromolecules like DNA, RNA, lipid, and protein inside the cell and eventually causes cell death. Persons who chronically exposed to UV radiation due to occupational or environmental exposure have a potential risk to develop cancer of the skin and internal organs. Polyphenols have a promising role in scavenging reactive oxygen species developed in our body due to a variety of exposure including radiation. In the present study, we are therefore evaluating the role of sinapic acid (SA), a polyphenol, in scavenging UV radiation-induced generation of reactive oxygen species and eventual cell death. We have studied the biological activity of SA as a potential antioxidant on human peripheral blood mononuclear cells taken from human volunteers.
Methods : Graded concentration of SA in different solvent mediums was applied to UV exposed peripheral blood mononuclear cells for evaluating the best effective delivery system and concentration against UV radiation.
Result: After series of experiments, we have observed that even at 10 µM concentration, SA is effective against UV induced cellular death.
Conclusion: Finally, from this work we are inviting lots of works in this field to establish this molecule as a medicine to reduce radiation-induced cellular death
Keywords
Sinapic Acid, UVB Radiation, Antioxidant, Cytotoxicity, Peripheral Blood Mononuclear Cell.
Introduction
Ultraviolet radiation is a potent environmental carcinogen, induces oxidative and inflammatory skin damage and eventually cancer. Naturally occurring polyphenols are potent antioxidants and an- ti-inflammatory agents which can potentially reduce the damaging effects of ultraviolet (UV) rays on the human body. Reduction in the ozone layer in the atmosphere promotes considerable UV ra- diation to enter into the earth surface which may cause damages to the biological systems. Although both UVA and UVB are po- tentially harmful, the effect of UVB is more severe as it causes burn to the absorbing tissues. UV ray can penetrate the skin and contributes its damaging effects mostly on the keratinocyte and fibroblast. Most of the UVB is absorbed in the epidermis layer of the skin leaving little to enter into the dermis layer.
UVB induced damages most precisely causes cell death and apop- tosis although not known to us that whether it may induce cell necrosis or not. Exposure to UVB causes formation of reactive ox- ygen species (ROS), cell cycle arrest, activation of various genes and cell markers [1]. Among all three types of UV rays, UVA and UVB rays can penetrate in human body. The UVA rays possessing the longest wavelength can reach deeper into the human skin, be- neath the dermis layer, while, comparatively little UVB rays can penetrate into the dermis layer. UVC rays are fully absorbed by ozone layer and cannot reach the Earth surface. UVB is potential- ly the more dangerous than UVA rays and it causes burning of the cells and responsible for producing skin cancer.
The carcinogenicity of UVB is well established and has been un- derstood as a potential damaging agent to DNA from which gene mutation arises. On the other hand, UVA is generally far more abundant on earth surface, and goes deeper into the skin surface, and therefore contribute equally to the carcinogenicity of sunlight. In contrast to UVB, UVA is hardly absorbed by DNA and there- fore less harmful from UVB regarding carcinogenic potential [2]. Therefore, whenever the UVB rays hit any soft tissue it causes cell death or alteration in cell cycle. The intensity of the UV exposure depends on the geographical position of a particular area. In trop- ics, the solar UV is more intense than in higher latitudes and the intensity of exposure decreases in arctic and polar region.
That’s why people in equatorial and tropical region are more vul- nerable to UV exposure than any higher latitudes. But the question is not only limited to solar exposure of UV rays. People are ex- posed to UVB rays nowadays through occupational, medical and cosmetic exposure from UV. Sun beds are extensively used in tan- ning parlous to tan skin and it uses UVB. Regular uses of Sun beds may contribute significantly to person’s annual cumulative UVB exposure. UV sources are used in various diagnostic and therapeu- tic purposes, and intensity may vary according to the type of treat- ment. Arc welding, UV light manufacturing industry, use of UV in research are also the potential contributing factors for UV ex- posure. All these route of exposure either alone or in combination give a high cumulative exposure to people working in these fields. Therefore, occupational exposure and environmental exposure can lead to a considerable risk from UV exposure on human body.
Polyphenols possess diverse functions including antioxidant, an- ti-inflammatory, antimutagenic and anticancer activities. To ex- plore the role of polyphenol in amelioration of UVB induced cell damage we planned to work out the effect of sinapic acid (SA) on UVB exposed peripheral blood mononuclear cells (PBMC). The polyphenols, with over 8000 currently known compounds, repre- sent one of the largest chemical groups in the plant kingdom, clas- sified into 4 major classes (phenolic acids, flavonoids, stilbenes, and lignans) [3].
SA is one of the four most common hydroxycinnamic acids and is widespread in the plant kingdom. It has been proposed as a po- tent antioxidant by many researchers, [4-6]. In recent years SA and some of its derivatives (sinapine, 4-vinylsyringol, sinapoyl esters, syringaldehyde) have gained tremendous attention because of their various biological activities as an antimicrobial, anti-inflammato- ry, anti-cancer, and anti-anxiety activity. It has also a potent an- ticholinesterase activity and thereby can be used in diseases like Myasthenia Gravis and other neurological disorders like Alzhei- mer’s disease, Parkinson’s disease, etc. [7]. SA, like other phenolic compounds, can be found in fruits, grains, and vegetables as well as in some medicinal plants and species (for instance borage, sage, mace, or rosemary) [8].
The protective properties of exogenous antioxidants have been documented in a number of epidemiological, interventional and in vitro studies [9]. Therefore, we hypothesized that SA could work to ameliorate radiation induced cellular toxicity. To evaluate the role of SA for combating UV radiation-induced damage, we have studied its effect on UVB exposed cells. Further, in this study, the solvent for SA has been studied to evaluate the best solvent for its delivery to get the maximum effectiveness.
The effect of drugs and its efficacy depends on the density of bio- available molecules in the system and its half-life [10]. An increase in bioavailable molecules in the blood increases the concentration of the said drug at the target site. Therapeutic drugs are generally low-molecular-weight molecules that result in non-specific dis- tribution within the body. This helps it to excrete itself through renal filtration resulting in rapid renal clearance and concomitant short plasma circulatory time [11, 12]. Although polyphenols are a good choice for combating diseases, dietary supplementation of flavonoids cannot reach the therapeutic concentration at the target site because of their low absorption rate from the gastrointestinal tract and low bioavailability [13]. The usual and best possible way of any drug administration is the oral route [13]. Polyphenols are poorly absorbed from the gut [13, 14] and therefore the alternative approach of administration may be required to raise the therapeu- tic concentration at the target site. Therefore, in this study we have also sought the alternative route of flavonoid delivery and selec- tive organic solvent for SA delivery.
Small nonprotein molecules with electrophilic properties are trans- ported through the blood to their target site by binding with albu- min. It is efficient for its relatively long half-life and abundance [14, 15]. Regarding its binding and transporting activity, it is im- portant to find out it’s mode of interaction with a drug molecule. Polyphenols are small electrophilic molecule and nonprotein in nature and therefore transported inside the plasma with the help of albumin through binding. We are therefore interested to study the interaction of SA with albumin to compare the bioavailability of SA and its biological activity in different solvent systems.
We have chosen three solvents: SA dissolved in alcohol, SA dis- solved in dimethyl sulfoxide (DMSO) and SA dissolved in 13% PEG-4000 as stock solutions for solvents. The binding constant of SA in these three solvents with albumin has been judged cru- cially to follow the extent of SA delivery to the biological system and carrying of SA in the system by binding with albumin. The stock solution for PEG, alcohol and DMSO were 20%. 14µl of SA (0.00224 gm SA in 2 ml of stock solution, i.e, 400µl alcohol in 1600 µl H2O) in 20% alcohol was dissolved in 1986 µl of PBS to meet the final concentration as 0.01%, which is then used to incu- bate cells. Similarly, the final concentration of DMSO is 0.005% and PEG is 0.01% in working solution in which the peripheral blood mononuclear cells were cultured and incubated for further study.
To evaluate the biological activity and efficacy of candidate sol- vent for transporting SA into the blood and its bioavailability at the blood and target site we have done in vitro study on human PBMC.
Here we studied the effects of UVB on PBMC to show the cyto- toxicity which may also true for keratinocytes. We used cultured PBMC as study model because it is easy to culture and maintain. In the later stage of our study we studied the effect of UVB expo- sure with and without SA on immortalized keratinocytes (HaCat).
Materials and Methods
Subject Selection
For in-vitro experiments 5ml of EDTA anticoagulated whole blood were taken from 6 healthy male volunteers. The average age was 34 years with 65 kg average weight. All the subjects were nonsmoker and neither has any known disease or nor under any medication. Their average blood pressure and heart rate were 124/86mmHg and 80 beats/min. Written inform consent were taken from each volunteers before enlisting their name in the study. From each participant cells were separated and studied in triplicate for each experiment. Therefore, in each experiment like, MTT assay, LDH assay, Nitric oxide assay, plasma free sialic assay the number of observation were 18 for each concentration and solvent of SA.
Reagents and Materials
SA (≥98%, powder), and phosphate buffer solution (PBS, pH 7.4) were purchased from Sigma-Aldrich, USA and used without fur- ther purification. PEG-4000, ethyl alcohol, and DMSO were pur- chased from Merck Millipore. All the solutions were prepared us- ing 0.1 M of PBS buffer at pH 7.4. LDH - cell cytotoxicity assay kit was purchased from HIMEDIA and kept at -200C until work. MTT was purchased from SRL, dissolved in 0.1 M PBS freshly and kept in dark until work. Ficoll-Histopaque and phytohaemo- agglutinin were purchased from Sigma. Immortalized keratinocyte cell line ‘HaCat’ was obtained from Thermo Fisher Scientific.
Instruments
All the UV-vis spectra were recorded on Hitachi U-4100 spectro- photometer at 298 K. The measurements at the respective wave- lengths were taken using photometry option in the instrument.
The emission spectra were measured on F-7000 Hitachi Spectro- fluorimeter equipped with a 1.0 cm path-length quartz cell and a circulating water bath, using an excitation wavelength of 280 nm. All the measurements were done at the micromolar range to avoid the aggregation and inner filter effect.
Sample Collection and Preparation
On the day of experiment peripheral blood was collected from healthy volunteers by venipuncture with EDTA-anticoagulated auto- claved polypropylene tube. From whole blood plasma cell compart- ments were separated using centrifugation. PBMCs were separated from the whole blood by the usual method of ficol density gradient centrifugation. Written informed consent was taken from every vol- unteer before taking their blood. Ethical principles followed by the institute are guided by rules as formulated by the Indian Council of Medical Research and these are in agreement with Helsinki rules.
Isolation and Irradiation of Peripheral Blood Mononu- clear Cells
Whole blood was collected from healthy human volunteers by phlebotomy. EDTA anti-coagulated whole blood was then subject- ed to ficol density gradient for isolation of PBMCs. Buffy coat was isolated by centrifugation and PBMCs were then washed in cold PBS and incubated at 37ºC for 1 hour with 5% CO2 in a CO2 incu- bator. Cells were then subjected to UVB after 15 min incubation under 37ºC with different concentration of SA dissolved (DMSO, alcohol, and PEG) in different solvents according to the design of the experiments in a CO2 incubator. The viability of cells was evaluated by MTT assay before and after SA administration with UVB exposure.
The UVB source consisted of a bank of four Philips TL-12 lamps (Philips, Eindhoven, Netherland). These lamps emit UV rays over the range of 250-400 nm, primarily in the UVB region (280-320 nm), with a peak at 315 nm. PBMC suspended in PBS in Petri dishes (2 x 106 cells in 2 ml per dish), were placed under the TL- 12 lamps after removal of the lid, and exposed to single doses of UVB (selected range of radiation was 10 mJ/cm2) (Becton Dickin- son) [16]. Before use, the culture media of the petri dishes (O 35 mm; Greiner) were treated with a graded concentration of SA (1 µM, 5 µM, 10 µM, 20 µM, 35 µM, 40 µM, 50 µM) in three dif- ferent solvent mediums. For each medium and each concentration of SA, MTT assay was performed to evaluate the percentage of viable cells present with and without administration of SA to UVB exposure. For each of the biological test 1µM was the lowest con- centration of SA (data not shown).
MTT Assay
From each subjects three aliquots of PBMC prepared were sub- jected to MTT assay. MTT assay was carried out on UVB-ex- posed PBMCs to evaluate the effect of SA in ameliorating UV radiation-induced cell death. SA was administered through three different solvent systems to compare its efficacy in the solvent systems. In brief, the single-cell suspension was made from fresh human blood by ficol. Cells were then washed in cold PBS and seeded in 96 well plates to incubate first at 37ºC for 1 hour with 5% CO2 in a CO2 incubator with SA dissolved in different sol- vent media (in alcohol, in PEG, and in DMSO). Briefly, 1 x104 to 5 x104 human PBMCs were treated with SA in three particular solvent systems prior to incubating under UVB rays (10mJ/cm2). The graded concentration of SA was also used in a similar design but a different set of experiment was performed to select the best effective concentration of SA against radiation-induced cellular toxicity. Cells were then incubated with 50 µl of MTT solution for 2.5 hrs at 370C. Formazan crystals formed, was then dissolved in 150 µl MTT solvent (4 mM HCl, 0.1% NP40 in isopropanol) and the absorbance was measured at 590nm. The effect of SA on radiation-induced cellular toxicity after UVB was compared to the cytotoxicity developed (if any) in control untreated cells, assigned here as 100%. Negative control was prepared by addition of MTT in cell-free PBS. Samples were prepared for each concentration and each solvent of SA in triplicate. Controls were prepared in same fashion without the exposure of SA and UVB.
Cytotoxicity was determined by corrected absorbance: % cytotoxicity = 100x (control-sample) Cell Cycle Analysis
SA treated (with 5 µM, 10 µM, 20 µM, 30 µM, 50 µM of SA) and untreated PBMC cells after washing with PBS were plated in a 24-well plate at a density of 1x106 cells/well and exposed to UV B irradiation at 10 mJ/cm2. Following incubation for 2 h, cells were harvested and re-suspended in PBS and fixed in 70% etha- nol at 40C overnight. They were then washed twice in cold PBS and incubated with propidium iodide staining solution (Beyotime Institute of Biotechnology, Haimen, China) for 30 min at room temperature. The percentage of cells at various phases of the cell cycle, namely the G0/G1, S and G2/M phases, were determined by flow-cytometric analysis of 1x106 cells. Same experiment was conducted with cultured immortalized keratinocytes, HaCat. In case of keratinocytes the dose of UVB were graded in one experi- ment to determine the dose dependent inhibition of cell cycle and in other the graded doses of SA were prior to UVB radiation and culture.
Assay of Cell Necrosis
Cytoplasmic leakage of LDH in extracellular space and measure- ment of this LDH is an indirect way to access the extent of cellular necrosis. Therefore to study the extent of necrosis PBMCs were separated from whole blood by ficol and then harvested in RPMI 1640 medium (HiMedia) using Dulbecco’s modified Eagle medi- um (HiMedia) and 5% fetal bovine serum and 5% CO2 in a 96 well microtiter plate at a density of 1x104 - 5x104 cells per well. The cells were incubated overnight at 37º C equilibrated with 5% CO2. After completion of the culture, lysis solution was added to each well and incubated further for another 45 minutes at the same condition. The contents were then centrifuged at 2000 rpm for 5 minutes and the supernatant was restored in a new vial. To the supernatant, LDH assay buffer was added and subjected to incu- bation in dark at 37ºC for about 25 min and the absorbance was taken at 520nm. Percentage of cytotoxic cells was calculated by supernatant, LDH assay buffer was added and subjected to incu- bation in dark at 37ºC for about 25 min and the absorbance was taken at 520nm. Percentage of cytotoxic cells was calculated by supernatant, LDH assay buffer was added and subjected to incu- bation in dark at 37ºC for about 25 min and the absorbance was taken at 520nm. Percentage of cytotoxic cells was calculated by the following formula according to Chan et al. [17] with mini- mum modification. Samples were prepared for each concentration and each solvent of SA in triplicate. Controls or untreated cells were prepared neither with SA nor with UVB. Negative controls are prepared in same way without addition of cells according to Chan et al. [17].
% Cytotoxicity = 100x (Corrected reading from test well - Cor- rected reading from untreated well)/(Corrected reading from un- treated well- corrected maximum LDH release negative control).
Sialic Acid
Assay Plsama free sialic acid was estimated from every sample (whole blood) exposed to UVB with and without SA administration by the method of Jourdian et al. [18].
Nitric Oxide Synthase Activity Assay
Nitric oxide synthase (NOS) activity was measured by Nitric ox- ide assay kit (colorimetric) from Abcam (ab211083). Briefly, PB- MCs were cultured with and without SA prior to UVB exposure for 15 min. Then, they were washed twice with ice-cold PBS and then homogenized in homogenization buffer (250mM Tris EDTA, 10mM EDTA, 10mM EGTA). Cellular debris and unbroken cells were precipitated by centrifugation at 12000 rpm for 15 min and the supernatant was used for NOS activity study. The supernatant was subjected to react with Griess reagent 1 and 2 (provided with the kit) and the colour generated was studied for its intensity colo- rimetrically at 540nm.
Serum Amyloid A Assay
The PBMCs were cultured on Poly-D Lysine/Laminin coated plates. The cells were then irradiated under UVB source with and without graded concentration of SA. PBMCs were then harvested and total protein extracted for ELISA assay to detect the serum amyloid-A level using commercially available α- amyloid kit (Bio- Assay Systems, Hayward, CA).
Assay of ROS Scavenging Activity
a) DPPH assay
The DPPH (2, 2-diphenyl-1-picrylhydrazyl) radical scavenging activity was determined according to the method of Xie et al [19]. The initial stock solution of SA was prepared in both alcohol-water and DMSO medium. The required concentration for the assay (1, 5, 10, 20, 30, and 50µM) was then achieved by further dilution of the stock solutions with double distilled water. According to the method, 0.5 mL of the sample solution with different concen- tration of SA was added to 3 mL of DPPH (0.1 mM) in methanol and mixed well. The solutions were kept in dark for 20 minutes and then absorbance was taken at 517 nm. The below-mentioned equation was utilized to calculate the percentage of scavenging activity:
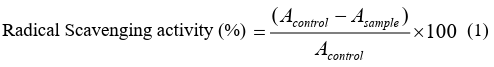
b) ABTS Assay
ABTS assay measure the relative ability of an antioxidant to scav- enge the ABTS generated in the aqueous phase, as compared with Trolox (water-soluble vitamin E analog) standard. The ABTS is generated by reacting with a strong oxidizing agent with the ABTS salt. It is a decolouration assay applicable to both lipophilic and hydrophilic antioxidants. The assay was performed according to the protocol of Re et al. [20] with different concentration of SA (1, 5,10, 20, 30 and 50µM), in which radical monocation of ABTS.+ (2,2’ azinobis-3 ethylbenzothiazoline-6-sulfonic acid) is generated by oxidation of ABTS with potassium persulfate and is reduced in the presence of such hydrogen donating antioxidants. This assay was also performed in two different solvent system of SA, alcohol water and DMSO water.
Statistical Analysis
Significance of differences between the mean of different param- eters has been studied by paired two-tail t-test and oneway ANO- VA to find out the difference between group means. Significance
Results
Steady-state fluorescence studies
The characteristics of the interaction of plasma protein with SA were studied initially in three different solvent media (DMSO/buf- fer, alcohol/buffer, PEG/buffer) using steady-state fluorescence. The plasma shows intrinsic fluorescence due to the presence of different plasma proteins mainly serum albumin. Serum albumin of difference was studied against a particular concentration of SA treated UVB exposed group and SA untreated UVB exposed group. The computed F score for ANOVA was compared with crit- ical F to find out the level of significance [21].
contains various fluorophores such as tryptophan (Trp), tyrosine (Tyr) and phenylalanine (Phe) residues, among them Trp residue shows the major while Tyr shows minor contribution for intrinsic fluorescence of serum albumin [21, 22]. Hence, we have moni- tored the fluorescence spectra of the blood plasma by exciting at 280 nm where both the Trp and Tyr residues contribute to the emis- sion spectra (vide Figures 1a, 1b).
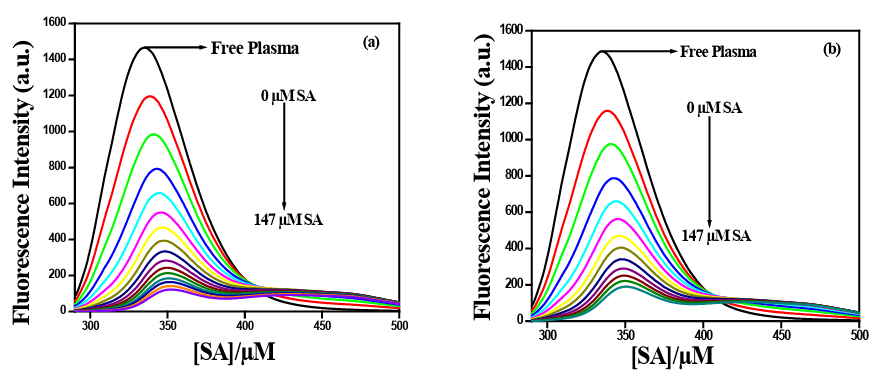
Figure 1: Fluorescence quenching spectra of plasma protein (albumin) in the presence of varying concentrations of SA at 298 K; λex= 280 nm, pH = 7.4 (0.1N PBS). Excitation band pass =5 nm and Emission band pass = 5 nm. (a) alcohol/buffer medium; (b) DMSO/buffer medium.
Evaluation of Binding Constant To understand the interaction of SA in different solvent medium with human plasma, we have exploited the fluorescence titration data using the following equation (equation 2) [6, 23-25].
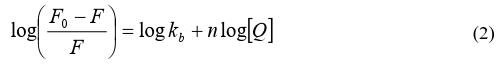
where F0 and F are the fluorescence intensities in the absence and presence of quencher molecule (Q), respectively, kb is the binding constant, [Q] is the concentration of the quencher and n is a num- ber of binding sites.
The binding constant of SA with human plasma decreases 10 times in PEG-buffer medium (kb = 2.344x104 M-1) in comparison to DMSO and alcohol, while that of DMSO/buffer and alcohol/buffer remained almost comparable (vide Figure 2, table 1).
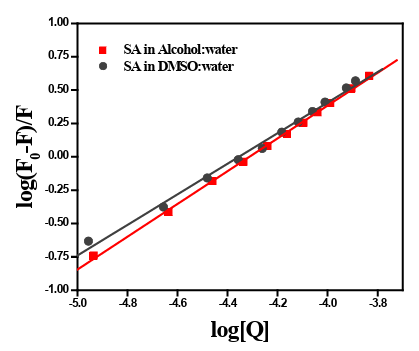
Figure 2: The double-logarithm plot of log[(F0-F)/F] vs. log[Q] for the interaction between human blood plasma with SA in two different media.
Table 1: List of binding constant (kb) and binding sites (n) for the interaction of SA in different media with human blood plasmaat 298K.
|
|
Medium |
kb (M ) -1 |
n |
R2 |
|
Human Blood Plasma |
Alcohol-buffer |
2x105 |
1.22 |
0.99 |
|
DMSO-buffer |
1x105 |
1.14 |
0.99 |
Figure 1a and 1b depict the effect of gradual addition of SA in al- cohol/buffer and DMSO/buffer respectively to human blood plas- ma. The percentage of alcohol and DMSO and PEG in working conditions was less than 0.01 %. The intrinsic fluorescence of the blood plasma decreases with the gradual addition of SA in both the medium, which indirectly indicated that, the increase in the bind- ing of SA with plasma protein and formation of a non-fluorescent complex. As evident from the plot, with increasing SA concentra- tion, gradual quenching of intrinsic fluorescence of plasma protein was observed in both alcohol/buffer and DMSO/buffer media.
Based on the above results we have performed the biological as- says by dissolving SA in all of the above-mentioned media.
Result of MTT assay, Sialic acid, LDH-cytotoxicity, ni- tric oxide synthase, serum amyloid A assay
The efficacy of SA to ameliorate radiation-induced cellular dam- age has been studied by evaluation of percentage of cytotoxicity and viable cell on UVB exposed PBMCs in three different solvent systems (DMSO, Alcohol, PEG). Percentage of viable cells and cytotoxicity were evaluated by MTT assay and LDH-cytotoxic- ity cell necrosis assay respectively. Cell death by tissue necrosis has been studied by nitric oxide synthase activity. UVB and SA exposed cultured PBMCs extract has been studied and compared for Serum amyloid-A assay in cells before and after radiation ex- posure. MTT assay on PBMCs after radiation exposure have been studied and showed that the prior administration of SA to PBMCs before UVB exposure is beneficial for cells as they survive more after treatment with SA. A significantly higher percentage of vi- able cells (p<0.05) were observed in aliquots incubated with SA, when compared between without exposure of SA and with grad- ed concentration of SA in UVB exposed cells dissolved in both DMSO and alcohol. No dose-response pattern has been seen in our study considering all the concentration of SA together. However, up to 10µM of SA concentration the percentage of viable cell in- creases in a dose response pattern. The percentage of the viable cell after higher dose (above 20µM) of SA administration with UVB rather showed a non-dose responsive status. We have studied each concentration of SA in triplicate with a minimum concentration of 1µM (data not shown) and found a dose response up to 10µM of SA. The number of viable cell in case of alcohol/buffer and DMSO /buffer system was highest at a concentration of 10µM (61.74%) and lowest at 1µM (41.36%) which shows a dose response rela- tionship. At the concentration 35, 20 and 5 µM the percentage of cell viability were 52, 48 and 49 respectively. Similar results were also obtained in DMSO and PEG buffer system (vide Figure 3).
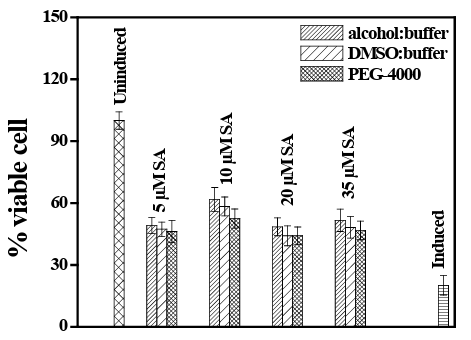
Figure 3: Comparative analysis of percent viable cells (analyzed by MTT assay) after administration of different concentrations of SA in three different solvent systems, on UVB exposed human pe- ripheral blood mononuclear cells.
Assay of cell cycle analysis
Result indicate a fall in cell growth and division as it exposed to UVB rays in both normal PBMC and in HaCat cells. Co-adminis- tration of SA reduces the percentage of cell death and restore some cell division and growth which is maximum in 30µM of SA admin- istration in both PBMC and HaCat cells. The percentage of apop- totic cell death and necrotic cell death are highest at UVB exposure without any co-administration of SA. Apoptotic and necrotic cell death was reduced efficiently after SA administration in the culture medium before exposed to UVB radiation. However, graded con- centration of SA can successfully increase the percentage of live cell after UVB exposure and this showed a dose response curve up to 30µM concentration. Beyond this level, the percentage of live cell reduced again instead of higher concentration of SA applica- tion. Again, the percentage of apoptotic and necrotic cells showed higher concentration when the culture was maintained without SA or very low concentration of SA. Both in PBMC and in HaCat the percentage of necrotic cell is highest in UVB exposure with only 10µM of SA. Whereas, the percentage of apoptotic cell is almost similar in UVB exposure without SA and with 5 µM of SA when maintained in PBMC. The cultured HaCat shows slightly different concentration of apoptotic cell when maintained with 5 µM of SA with UVB in comparison to only UVB.
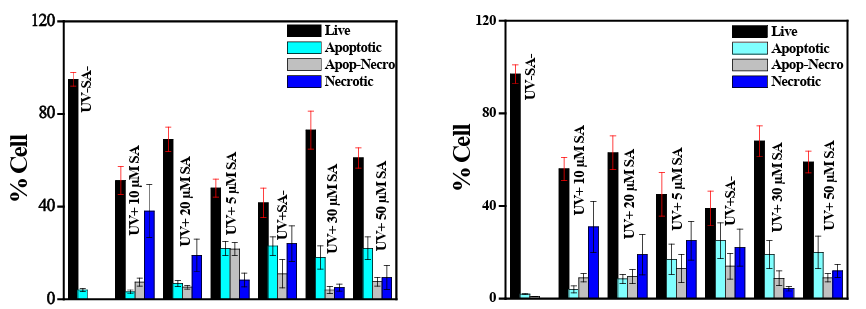
Figure 4 a: PBMC cell culture with UVB and SA Figure 4b: HaCat culture with UVB and SA
Figure 4: Assay of Cell growth and cell death by flow cytometric analysis using human blood peripheral momonuclear cells and immor- talized keratinocytes (HaCat) after UVB exposure with and without SA administration.
LDH assay
LDH level is an indicative of cellular death. The percentage of cyto- toxicity induced by UVB and its amelioration by co-administration of SA showed interesting results. The percentage of cytotoxicity in case of SA administered through alcohol/buffer system was in the order of 5µM (38.49%)>20µM (37.92%)>10µM (35.78%)>35µM (20.68%) and that for DMSO/buffer media was 20µM (53.15%)>5µM (45.10%)>10µM (38.18%)>35µM (20.15%) and in case of SA in PEG medium that result was 5µM (58.03%)>10µM (55.69%)>20µM (52.33%)>35µM (43.00%). The observation primarily indicated that a concentration of 35 µM of SA was able best to prevent cell damage against UVB. Interestingly, the optimum concentration for alcohol/ buffer, DMSO/buffer and SA in PEG/buffer medium was 35µM. The percent cytotoxicity for alcohol/buffer, DMSO/buffer and PEG /buffer media was also comparable to each other (vide Figure 5). We have also studied the experiment with 1µM SA which showed a poor ame- lioration from UVB induced damage. The level of LDH before and after administration of SA in UVB exposed PBMC was significantly different (p<0.01) between 5 and 35µM of SA (p<0.05).
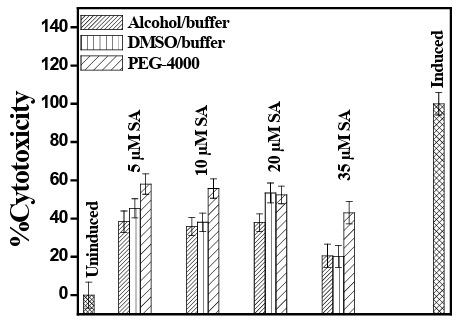
Figure 5: LDH Assay (mean ± SD) for % cytotoxicity of human PBMC exposed to UVB after prior administration of SA in DMSO/ buffer, alcohol/buffer media and PEG 4000 in buffer medium.
From sialic acid assay it has been shown that plasma free sialic acid concentration increases in UVB irradiated blood cells in comparison to blood cells irradiated under UVB with SA. Plasma-free sialic acid lev- el in irradiated blood cells is significantly higher (p<0.01) in compar- ison to un-irradiated samples. When human blood samples were sub- jected to UVB irradiation with a graded concentration of SA it showed a decrease in plasma-free sialic acid concentration. A concentration of 10 µM of SA has shown maximum effectiveness against UVB ir- radiation in this assay. The difference in sialic acid concentration be- tween UVB irradiated 10µM SA added cells and without SA added cells was also significant (p<0.05). The analysis indicated that alcohol/ buffer medium was the suitable one and the order of effectiveness was 10µM<5µM<1µM<35µM<20µM. In the case of the DMSO/buffer solvent system and SA-PEG, the order followed a similar trend as 10µM<5µM<1µM<35µM<20µM. In all three cases, the absorbance value was lowest at a 10µM concentration of SA, which suggested a lesser release of sialic acid from irradiated blood cell at that particular concentration (vide Figure 6).
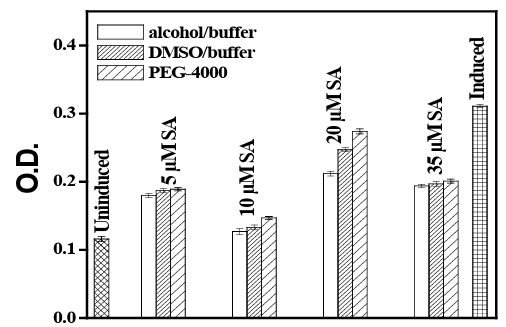
Figure 6: Sialic Acid Assay for evaluation of cell death (O.D630nm) of human PBMC exposed to UVB with prior administration of SA in DMSO/buffer and alcohol/buffer media and SA in PEG/ buffer medi- um.
Free radical scavenging activity of SA has been studied by DPPH and ABTS assay. It is seen from the result of DPPH activity that gradual increase in SA concentration increases the free radical scavenging activity of SA in a dose-dependent manner which is also significant (p<0.05). A similar result has also been observed when the ABTS assay performed to evaluate the Trolox equivalent (Figure 7). This study also showed a dose-response pattern when SA concentration was increased gradually.
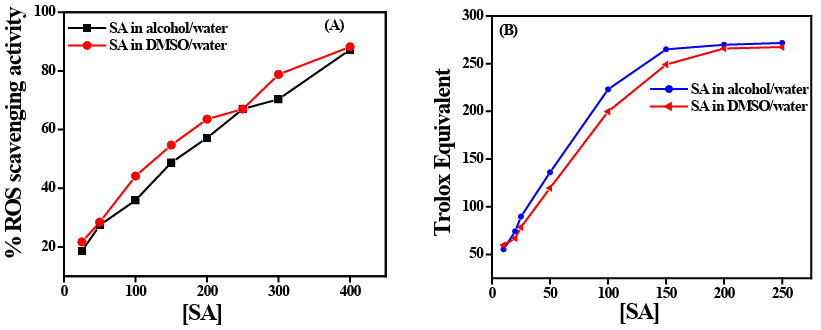
Figure 7: (A) ROS scavenging activity of SA before and after exposure of UVB measured by DPPH activity. (B) ROS scavenging activity of SA measured by Trolox equivalent through ABTS activity after UVB exposure.
Serum amyloid A (SAA) is an apolipoprotein and a major acute phase reactant which has a central role in the inflammatory re- sponse. SAA is expressed primarily in the liver, although it is also found in extrahepatic sources such as adipocytes and macrophages. SAA is involved in cholesterol sequestering and lipid metabolism and has been shown to induce extracellular-matrix-degrading en- zymes, pro-inflammatory cytokines and to recruit immune cells to sites of inflammation by chemotaxis [26]. Radiation can induce SAA in rodents and other primates [26]. Here we have shown that UVB induces SAA at a significantly high level (p<0.05) in com- parison to normal unexposed PBMCs. Radiation-induced SAA ex- pression is increased to show a dose-response relationship (data not shown here, when gradually the time of exposure has been in- creased from 5 min to 25 min). 15 min UVB exposure with graded concentrations of SA can reduce the SAA concentration in com- parison to UVB exposure without SA, although did not attain the normal unexposed level (Figure 8).
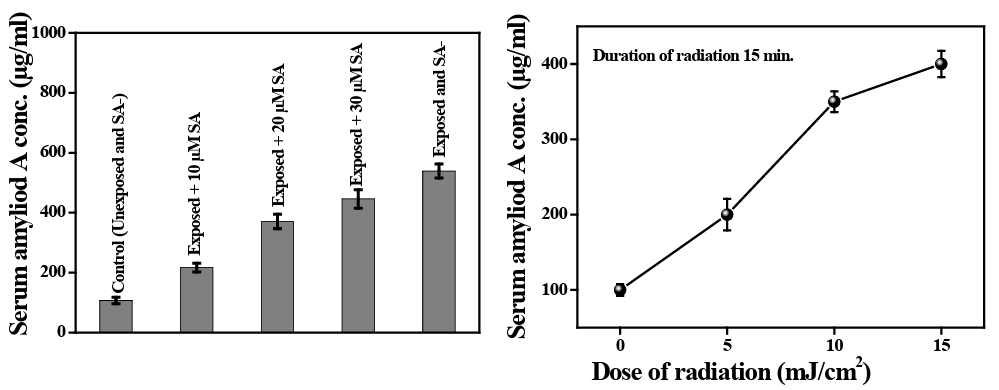
Figure 8: Serum amyloid A (SAA) level in UVB exposed human peripheral blood before and after administration of sinapic acid. The bar diagram shows the mean ±SD value of SAA before and after SA administration on UVB exposed blood cells.
Our experiment on nitric oxide synthase (NOS) activity after UVB exposure to PBMC revealed that the activity of NOS is increased dramatically when exposed to radiation and decreased when the cells are cultured with different concentration of SA. The nitrate concentration (due to NOS activity) is increased 4 fold after UVB exposure without administration of SA. A significant decrease (p<0.05) in nitrate concentration has been noted after graded con- centrations (1 µM, 5 µM, 10 µM, 20 µM, 35 µM, and 50 µM) of SA administration prior to UVB exposure in culture medium of PBMC cells. The lowest concentration of nitrate has been noted in cells treated with 35 µM of SA (vide Figure 9).
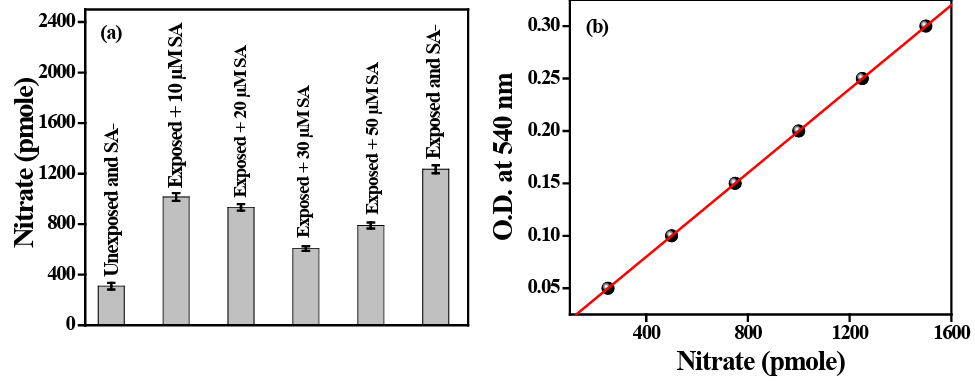 Figure 9: 9a represents the pmole concentration of nitrate evolved against OD level at 540nm. 9b represents the concentration of nitrate evolved in cells after UVB exposure with and without SA administration.
Figure 9: 9a represents the pmole concentration of nitrate evolved against OD level at 540nm. 9b represents the concentration of nitrate evolved in cells after UVB exposure with and without SA administration.
Discussion
UV radiation induces apoptosis through two biochemically and mor- phologically distinct processes, apoptosis and necrosis. Intricate cell biology shows that various cell lines undergo apoptosis following low doses of UVB irradiation, whereas prolonged exposure induces necrosis [19]. UVA and UVB both are potentially harmful and both can induce squamus cell carcinoma (SCC) in nude mice but the abil- ity to produce SCC of UVA alone is about 5000 to 10000 times low- er than UVB alone [27- 29]. Both in vitro and in vivo experiments and epidemiological studies indicate that long lasting chronic expo- sure to UVB is the main cause of SCC of the skin [27]. Before 1990, for this reason, only UVB was considered as carcinogenic, not UVA. It was revealed experimentally, that the UVA is less toxic than UVB for induction of melanocyte tumors in Monodelphis domestica [30]. Experiments with UVA on new born rodents and on isolated human skin cell couldn’t reproduce the results obtained with UVB [31, 32]. All these experimental evidence suggest that similar dose of UVB is more dangerous than UVA to produce melanoma and SCC. Besides skin, our lips, conjunctiva and iris muscles are also can be exposed to UV radiation. Therefore, although UVB cannot penetrate deep beneath the skin, these soft tissues are under the threat of being ex- posed and damaged. This is one of the major reasons for choosing UVB to explore the cellular effects on it and the ameliorating action of SA on UVB induced cellular damage.
UV radiation affects the biological system through disruption of the cellular homeostasis by damaging the membrane and macromole- cules inside the cell including DNA, RNA, and protein. ROS are generated by UVA and UVB which are highly reactive and cause alteration of the molecular structure of DNA, lipids, and proteins [32, 33] through inhibiting DNA repair pathway [33, 34]. In our study, post UV exposure cellular toxicity profile and cellular dam- age was assessed by MTT, NOS and LDH assay which shows dif- ferential damage to the cell and causes cellular death. The scope of our present paper is limited and not focusing to the mechanism of UVB induced apoptosis or cellular death, so, we are briefing the present reviews and experimental data on the proposed mechanism of UVB induced cell death to focus the effect of UVB irradiation on peripheral blood mononuclear cells. It has been shown that SA can effectively block the DNA damage after UVB and has a cyto-protec- tive role against UV radiation [35]. Several studies indicate the role of SA as an efficient antioxidant, an anti-inflammatory, anticancer, anxiolytic and anti-neurodegenerative molecule [35]. Low doses of UVB (≤15 mJ/cm2) had been shown to induce caspase-dependent apoptosis, while a significant low dose of UV-B (35 mJ/cm2) caused apoptosis with altered morphology, slower DNA fragmentation, and poly(ADP-ribose) polymerase degradation accompanied by in- creased Bcl-2 and high doses of UVB (80 mJ/cm2) induced necrosis [36]. Again, UVB-irradiated lymphocytes showed an increased lev- el of peroxidative enzymes and lipid peroxidationwhich indicates disturbances in antioxidant defense [37]. UVB and gamma radia- tion induced cell damage and subsequent death and the cell protec- tive effect of several compounds like sesamol, ferulik acid, Caffeic acid, circumin had been studied by several researchers [37-41]. The weakness of these studies are not a single one of those could not effi- ciently point out the direction , medium and dose of those compound to effectively ameliorate the effect of radiation from living cells. We have shown that SA effectively reduces generation of ROS in post UVB exposed peripheral blood mononuclear cells. May be, due to its hydrogen or electron donation property it can act as a free radical scavenger and antioxidant molecule. In this paper, we are trying to evaluate not only the cyto-protective effect of SA but also the effec- tive dose of SA in isolated in-vitro cellular system at which, it most effectively scavenges the ROS generated by UVB and prevents cel- lular death from UVB exposure. The study also indicates the best suitable solvent for SA delivery.
The pharmacokinetic study revealed that serum albumin is respon- sible for the transportation of SA in the blood, due to its ability to combine with serum albumin. It is also evident from our study that SA after administered to the ex-vivo human plasma, combine with plasma protein to transport to the cellular system. Study with HSA and BSA revealed that SA bind with albumin and thus reduces the fluorescence (data not shown). The maximum reduction of fluores- cence intensity has been observed with 147µM of SA, when it is gradually added to the human plasma (Figure 1). Reduction in flu- orescence is indicative of the quenching of SA with plasma protein. But it is not known what amount of these transported SA is released to the tissue space or target site. It is rather more complicated to work out. Actually, the biological activity is the reflection of the internalized SA into the cell which ameliorates radiation-induced cellular death.
The binding constant for PEG-SA solvent system to plasma is about 100 times low in comparison to alcohol-SA and DMSO-SA solvent system. However, we do not know the interaction profile of PEG with SA. We presume that this solvent system is not equivalent to the PEGylation of small protein drug which has immense interest as a drug delivery system as it increases the half-life of the drug attached covalently with the PEG or encapsulated in PEG. There- fore, it is preferable to study the ameliorating action of SA through DMSO-SA or alcohol-SA solvent system. On the other hand, it is reflected from our experiment that SA dissolved in a solvent like DMSO and alcohol shows considerably higher binding capacity with plasma which can induce greater bioavailability of circulatory SA to the target tissue. It is hypothesized that greater the bioavail- ability, greater will be the ameliorating action.
Administration of graded concentration of SA on the PBMCs sub- jected to UVB causes a reduction in radiation-induced cellular dam- age significantly. Significant reduction in cellular death and an in- crease in viable cells in SA administered PBMCs subjected to UVB is actually the indicative of beneficial role of SA against UVB in- duced cellular death. Reduction in LDH and NOS activity reflected the capability of SA to protect cells from UVB induced necrosis and radiation induced damage. We have done all the experiments of SA with 1 to 50 µM of concentration. The reason for choosing this range of SA is that we have found a significant beneficial effect in this range. Beyond 50µM concentration the cells undergoes shrinkage (data not shown) which may indicate the early signs of cell death. At a concentration of 100µM of Sinapic acid about 50% of the cells undergoes membrane corrugation and death. Therefore we have planned to hold the concentration of SA up to 50µM when admin- istered prior to UVB. Radiation causes generation of different types of ROS in the biological system. Disruption of balance between its production and scavenging can cause cell death and break of mac- romolecules like DNA, RNA and proteins which in turn damage the cells. SA is a class of phenolic molecule which efficiently scavenge generated ROS in the body and thereby protect the system. It can scavenge O•, OH•, -OOH, ONOO- and other ROS generated in the body as a result of radiation [42]. Anti-mutagenic and anti-carcino- genic potential of SA had also been studied by different groups [43].
SAA is a biomarker for radiation exposure [43] and we have also shown from our work that UVB exposure to whole blood causes an increase in SAA level. However, the result showed that, with simul- taneous administration of SA and UVB, the SAA is reduced signifi- cantly. Sprull has shown that SA causes a significant reduction in SAA level in mice after receiving 1-8 Gy whole body or partial body radiation [44]. Cyto-protective effects of SA have been studied by Kim and Lim on UVB exposed human keratinocytes [45]. Amount of UVB induced DNA damage and ROS generation has been shown to be decreased after a time and concentration-dependent SA admin- istration prior to UVB exposure on keratinocytes [46]. The expres- sion of DNA repair gene is increased in a dose-dependent manner when SA is applied simultaneously with UVB on keratinocytes [47]. Although we have significant number of literature in respect to the beneficial role of SA, still, the dose dependence of SA and solvent system for its delivery has not been studied in radiation induced cy- totoxity. To our knowledge, we are the first to propose the effective solvent for SA delivery and its effective dose at which it can best ameliorate the radiation-induced cellular damage at least in vitro system. By this preliminary work we think, we are inviting lots of works in this field to evaluate the role of SA with its effective dose and solvent in radiation induced toxicity.
Conflict of Interest
No conflict of interest exist.
Acknowledgement
We are grateful to the Central Instrumental Facility of department of Chemistry and department of Biology, Presidency University for their constant support. The technical support for human cell cul- ture is provided by CSIR-Indian Institute of Chemical Biology and Jadavpur University. The financial support has been provided by Department of Science and Technology and Biotechnology, West Bengal, India.
Ethical Permission
Written informed consent for work with human blood has been ob- tained from every participant before taking their blood and Ethical permission has been obtained from Presidency University, Kolkata, by Human Ethical Committee, to carry on the work, which follows the Helsinki rule.
Consent to Publish
Every contributor of this work, and all of my coauthor do agree to publish this work.
Author’s Contribution
Adity Bose and Sarmishtha Chand contributed to the study concep- tion and design of the work. Material preparation, data collection and analysis was done by Prasenjit Mondal. The first draft was writ- ten by Sarmishtha Chanda and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Funding
Funding has been provided by West Bengal Department of Science and Technology and Biotechnology (WBDST; grant no: 105/DST/ AS/2017).
Competeing Interest
The authors have no relevant financial or non-financial interests to disclose.
Availability of data and materials
All of the raw data and materials can be submitted at any point of submission of this manuscript.
References
- Khalil, C., Shebabi, W. (2017) UVB damage onset and progres- sion 24h post exposure in human-derived skin cells. Toxicol Rep, 4: 441-449.
- de Gruijl, F.R. 2000: Photocarcinogenesis: UVA vs UVB. Methods Enzymol. 319: 359- 366.
- Pandey, K.B., Rizvi, S.I. (2009) Plant polyphenols as dietary antioxidants in human health and disease. Oxidative medicine and cellular longevity, 2: 270-278.
- Natella, F., Nardini, M., Di Felice, M et al. (1999) Benzoic and Cinnamic Acid Derivatives as Antioxidants: Structure-Activity Relation. Journal of agricultural and food chemistry, 47: 1453- 1459.
- Pekkarinen, S.S., Stockmann, H., Schwarz, K. et al (1999) An- tioxidant Activity and Partitioning of Phenolic Acids in Bulk and Emulsified Methyl Linoleate. Journal of Agricultural and Food Chemistry, 47: 3036-3043.
- Sengupta, P., Ganguly, A., Bose, A. (2018) A phenolic acid based colourimetric ‘naked-eye’chemosensor for the rapid de- tection of Cu (II) ions. Spectrochimica Acta Part A: Molecular and Biomolecular Spectroscopy, 198: 204-211.
- Niciforovic, N., Abramovic, H. (2014) Sinapic Acid and Its Derivatives: Natural Sources and Bioactivity. Comprehensive reviews in food science and food safety 13: 34-51.
- Zych, M., Kaczmarczyk-Sedlak, I., Wojnar, W. et al (2018) The Effects of Sinapic Acid on the Development of Metabolic Disorders Induced by Estrogen Deficiency in Rats. Oxidative medicine and cellular longevity, 9274246: 1-11.
- Kampa, M., Alexaki, V.I., Notas, G. et al (2004) Antiprolifera- tive and apoptotic effects of selective phenolic acids on T47D human breast cancer cells: potential mechanisms of action. Breast Cancer Research 6: 1-12.
- Larsen, M.T., Kuhlmann, M., Hvam, M.L. et al (2016) Albu- min-based drug delivery: harnessing nature to cure disease. Molecular and cellular therapies, 4:1-12.
- Markovsky, E., Baabur-Cohen, H., Eldar-Boock, A. et al (2012) Administration, distribution, metabolism and elimina- tion of polymer therapeutics. Journal of Controlled Release 161: 446-460.
- Sleep, D., Cameron, J., Evans, L.R. (2013) Albumin as a ver- satile platform for drug half-life extension. Biochimica et Bio- physica Acta (BBA)-General Subjects, 1830: 5526-5534.
- Wang, S., Zhang, J., Chen, M. et al (2013) Delivering flavo- noids into solid tumors using nanotechnologies. Expert opinion on drug delivery, 10: 1411a-1428.
- Yhee, J.Y., Song, S., Lee, S.J. et al (2015) Cancer-targeted MDR-1 siRNA delivery using self-cross-linked glycol chitosan nanoparticles to overcome drug resistance. Journal of Con- trolled Release, 198: 1-9.
- Vachali, P.P., Li, B., Besch, B.M. et al (2016) Protein-flavonoid interaction studies by a Taylor dispersion surface plasmon reso- nance (SPR) Technique: A novel method to assess biomolecular interactions. Biosensors, 6: 6-9.
- Teunissbn, M., Sylva-stehnland, R.M.R., Bos, J.D (1993) Ef- fect of low-dose ultraviolet-B radiation on the function of hu- man T lymphocytes in vitro. Clinical & Experimental Immu- nology 94: 208-213.
- Chan, A.W., Tetzlaff, J.M., Gotzsche, P.C. et al. (2013) SPIR- IT 2013 explanation and elaboration: guidance for protocols of clinical trials. Bmj, 346: 1-42.
- Jourdian, G.W., Dean, L., Roseman, S (1971) A periodate-re- sorcinol method for the quantitative estimation of free sialic acids and their glycosides. The j biol chem, 246: 430-435.
- Xie, J., Schaich, K.M. (2014) Re-evaluation of the 2,2-Diphe- nyl-1-picrylhydrazyl Free Radical (DPPH) Assay for Antiox- idant Activity. Journal of agricultural and food chemistry, 62: 4251-4260.
- Re, R., Pellegrini, N., Proteggente, A. et al. (1999) Antioxidant activity applying an improved ABTS radical cation decoloriza- tion assay. Free radical biology and medicine, 26: 1231-1237.
- Abou-Zied, O.K., Al-Shihi, O.I. (2008) Characterization of sub- domain IIA binding site of human serum albumin in its native, unfolded, and refolded states using small molecular probes. Journal of the American Chemical Society, 130: 10793-10801.
- Paul, B.K., Ganguly, A., Guchhait, N. (2014) Deciphering the interaction of a model transport protein with a prototypical im- idazolium room temperature ionic liquid: effect on the confor-mation and activity of the protein. Journal of Photochemistry and Photobiology B: Biology, 133: 99-107.
- Lakowicz, J.R. (1983) Principles of Fluorescence Spectrosco- py, Plenum Press, 3rd ed. New York.
- Bi, S., Ding, L., Tian, Y. et al (2004) Investigation of the inter- action between flavonoids and human serum albumin. Journal of Molecular Structure, 703: 37-45.
- Naveenraj, S., Anandan, S. (2013) Binding of serum albumins with bioactive substances–nanoparticles to drugs. Journal of Photochemistry and Photobiology C: Photochemistry Reviews, 14: 53-71.
- Sproull, M., Kramp, T., Tandle, A. et al. (2015) Serum amyloid A as a biomarker for radiation exposure. Radiation research 184: 14-23.
- De Laat, A., Van Der Leun, J.C., De Gruijl, F.R. (1997) Car- cinogenesis induced by UVA (365-nm) radiation: the dose-time dependence of tumor formation in hairless mice. Carcinogene- sis, 18: 1013-1020.
- Griffiths, H.R., Mistry, P., Herbert, K.E. et al. (1998) Molecu- lar and cellular effects of ultraviolet light-induced genotoxicity. Critical reviews in clinical laboratory sciences 35: 189-237.
- Ley, R.D. (2001) Dose Response for Ultraviolet Radiation A– induced Focal Melanocytic Hyperplasia and Nonmelanoma Skin Tumors in Monodelphis domestica. Photochemistry and photobiology, 73: 20-23.
- Robinson, E.S., Hill Jr, R.H., Kripke, M.L. et al (2000) The Monodelphis melanoma model: initial report on large ultravio- let A exposures of suckling young. Photochemistry and photo- biology, 71: 743-746.
- Berking, C., Takemoto, R., Satyamoorthy, K. et al (2004) In- duction of melanoma phenotypes in human skin by growth fac- tors and ultraviolet B. Cancer research, 64: 807-811.
- Kulms, D., Schwarz, T. (2000) Molecular mechanisms of UV- induced apoptosis. Photodermatology, Photoimmunology and Photomedicine: Review article 16: 195-201.
- Denning, M.F., Wang, Y., Tibudan, S. et al. (2002) Caspase ac- tivation and disruption of mitochondrial membrane potential during UV radiation-induced apoptosis of human keratinocytes requires activation of protein kinase C. Cell Death & Differen- tiation 9: 40-52.
- Lee, C.H., Wu, S.B., Hong, C.H. et al (2013) Molecular mech- anisms of UV-induced apoptosis and its effects on skin residen- tial cells: the implication in UV-based phototherapy. Interna- tional journal of molecular sciences, 14: 6414-6435.
- Chen, C. (2016) Sinapic acid and its derivatives as medicine in oxidative stress-induced diseases and aging. Oxidative medi- cine and cellular longevity, 357: 1614, 1-10.
- Caricchio, R., McPhie, L., Cohen, P.L (2003) Ultraviolet B Ra- diation-Induced Cell Death: Critical Role of Ultraviolet Dosein Inflammation and Lupus Autoantigen Redistribution. The Journal of Immunology, 171: 5778-5786.
- Prasad, N. R., Mahesh, T., Menon, V. P. et al. (2005) Photopro- tective effect of sesamol on UVB-radiation induced oxidative stress in human blood lymphocytes in vitro. Environmental tox- icology and pharmacology 20: 1-5.
- Srinivasan, M., Prasad, N. R., Menon, V. P (2006) Protective effect of curcumin on γ-radiation induced DNA damage and lipid peroxidation in cultured human lymphocytes. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, 611: 96-103.
- Prasad, N. R., Jeyanthimala, K., Ramachandran, S. (2009) Caf- feic acid modulates ultraviolet radiation-B induced oxidative damage in human blood lymphocytes. Journal of Photochemis- try and Photobiology B: Biology, 95: 196-203.
- Ramachandran, S., Prasad, N. R., Karthikeyan, S. (2010) Ses- amol inhibits UVB-induced ROS generation and subsequent oxidative damage in cultured human skin dermal fibroblasts. Archives of dermatological research, 302: 733-744.
- Prasad, N. R., Srinivasan, M., Pugalendi, K. V. et al. (2006) Protective effect of ferulic acid on γ-radiation-induced micro- nuclei, dicentric aberration and lipid peroxidation in human lymphocytes. Mutation Research/Genetic Toxicology and En- vironmental Mutagenesis, 603: 129-134.
- Kim, G.R., Lim, K.H. (2018) Cytoprotective effects of sinapic acid on human keratinocytes (HaCaT) against ultraviolet B. Biomedical Dermatology, 2: 1-7.
- Kale, A., Piskin, O., Bas, Y. et al (2018) Neuroprotective effects of Quercetin on radiation-induced brain injury in rats. Journal of radiation research, 59: 404-410.
- Assefa, Z., Garmyn, M., Vantieghem, A. et al (2003) Ultravio- let B radiation-induced apoptosis in human keratinocytes: cy- tosolic activation of procaspase-8 and the role of Bcl-2. FEBS letters, 540: 125-132.
- Zou, Y., Kim, A.R., Kim, J.E. et al (2002) Peroxynitrite Scav- enging Activity of Sinapic Acid (3, 5-Dimethoxy-4-hydroxy- cinnamic Acid) Isolated from Brassica juncea. Journal of agri- cultural and food chemistry, 50: 215884-5890.
- Ramachandran, S., Prasad, N. R. (2008) Effect of ursolic acid, a triterpenoid antioxidant, on ultraviolet-B radiation-induced cy- totoxicity, lipid peroxidation and DNA damage in human lym- phocytes. Chemico-biological interactions, 176: 99-107.
- Sengupta, P., Sardar, P.S., Roy, P. et al (2018) Investigation on the interaction of Rutin with serum albumins: Insights from spectroscopic and molecular docking techniques. Journal of Photochemistry and Photobiology B: Biology, 183: 101-110.