
International Journal of Botany and Horticulture Research(IJBHR)
ISSN: 2994-872X | DOI: 10.33140/IJBHR
Impact Factor: 0.98


Research Article - (2025) Volume 3, Issue 1
A Simple and Efficient Agrobacterium Tumefaciens-Mediated Transgenic System for Tetraploid Potato Cultivar Desiree
2International Potato Center (CIP), China Center for Asia-Pacific (CCCAP), China
3International Potato Center (CIP), Kenya
Received Date: Jul 18, 2025 / Accepted Date: Aug 15, 2025 / Published Date: Sep 17, 2025
Copyright: ©2025 Yan Feng and Jieping Li, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation: Xu, C., Wang, D., Qi, L., Chang, S., Yuan, P., et al. (2025). A Simple and Efficient Agrobacterium Tumefaciens-Mediated Transgenic System for Tetraploid Potato Cultivar Desiree. Int J Bot Hor Res, 3(1), 01-11.
Abstract
As the fourth staple food crop in the world, the potato has the potential to provide adequate amounts of energy, protein, and nutrients necessary for humanity. However, given the rapid expansion of the global population and the adverse impacts of climate change, there is an urgent need for enhanced yields and enduring resilience. This study addresses this need by presenting a simple and efficient transgenic system for the tetraploid potato cultivar Desiree. The system, built by testing different potato materials, explants types, selection medium, selection agents, and Agrobacterium tumefaciens EHA105 strain carried different vectors, stands out for its remarkable speed, producing positively transformed seedlings in about 40 days. Compared to some traditional methods with the pre-treatment of explants and two-step regeneration stages, which need more operations and time, in this system explants were directly inoculated with A. tumefaciens, a single-regeneration medium (SEG or SE) were used, which decreased operations and saved time. Leaf disc explants are eligible. The concentration of 20 μg/L is suitable for chlorsulfuron selection. The positive transgenic efficiency of tetraploid recipient Desiree is up to 68.79 %. Meanwhile, compare to diploid materials receptor CIP703541 and CIP705079, tetraploid materials Deisree have a lower ploidy change ratio after tissue culture process. This simple and fast transgenic system not only supports basic research on potato functional genes but also creates new potato materials for genetic engineering breeding, thereby contributing to the development of more resilient and productive potato varieties
Keywords
Potatoes, Transgenic System, Transgenic Efficiency, Desiree
Abbreviations
2,4-D: 2,4-Dichlorophenoxyacetic acid
IAA: Indole acetic acid
NAA: 1-Naphthaleneacetic acid
6-BA: 6-Benzylaminopurine
Introduction
Potato (Solanum tuberosum L.) is an important staple crop for human nutrition, ranking fourth globally, after rice, wheat and maize [1]. As the largest non-cereal food worldwide, potato is a perfect food, to provide humanity’s needs for energy, protein, and nutrition. To secure sustainable harvests, the best economic solution is to increase a staple crop’s resilience, yield and quality through breeding. One approach involves conventional breeding, harnessing genetic resources found in natural variation.
Alternatively, transgenic breeding integrates functional genes into the target genotype, precisely enhancing desired traits. Compared to traditional breeding, transgenic breeding is much more direct, precise, and time-efficient [2,3]. Certain successful transgenic methodologies in potato have employed protoplasts: the designated vectors were introduced into plant cells, and following selection, these cells harbored the target vectors, subsequently regenerating into new transgenic plants [4]. However, resulting from the need to remove the cell wall during the procedure, potato protoplasts are very weak. Further, the system concludes induced extraction and purification of protoplast, induction of callus from protoplast, roots regeneration from callus, means that need different mediums, which contain many chemical ingredients and the efficiency of positive transgenic events is low. Another method of introducing foreign DNA into cells is by using a propelled particle bombardment gene fragment deliver system, ‘gene gun’ [5]. However, this method produces high numbers of insert events at random sites and creates chimeras. The most stable and widely used method is transgenics via Agrobacterium tumefaciens. In potato, the first successful genetic modification using A. tumefaciens was accomplished in 1983 [6]. Since then, multiple research groups have dedicated significant efforts to improving the system’s efficiency by investigating the recipient plant’s conduciveness to transformation, explant establishment and growth, and improving growth mediums (REF). There are many successful cases, some high profile, of breeding companies using Agrobacterium-based transgenics to create new varieties. In 1995, Monsanto released the first biotech potato varieties developed from transformation via A. tumefaciens, NewLeafTM [7,8]. The new variety, derived from Russet Burbank, contained the crylIIA gene, which encodes the insect control protein Bacillus thuringiensis var. tenebrionis. More recently, Monsanto has developed NewLeaf PlusTM, a Russet Burbank variety with resistance to both Colorado Potato Beetle (CPB) and Potato Leafroll Virus (PLRV) [9]. J.R. Simplot Company successfully developed a low-acrylamide potato with black spot bruise resistance named InnateTM 1.0. However, the InnateTM 2.0 potato has the same traits as 1.0 traits with the addition of late blight resistance and improved cold storage capability. Furthermore, research performed at CIP used A. tumefaciens- mediated transformation to elevate late blight resistance levels of a few popular varieties by stacking three resistant genes. BASF developed Amflora, a high-starch potato that was approved in Europe, and Modena, and Fortuna with late blight resistance [10]. Further, the Russian Academy of Sciences developed insect resistance transgenic potato Elizaveta Plus and Lugovskoi Plus via Agrobacterium [11].
Although there are many successful transgenic systems for potato, a simple and efficient protocol will strongly support gene function research, especially for a traditional tetraploid such as Desiree, which always worked as the transgenic receipt model for economical tetraploid potato varieties. Several critical factors, such as the vector, A. tumefaciens strain, pre-culture period, explant type, culture mediums, and selection markers, will influence the time-cost and efficiency of potato transgenics [12]. The A. tumefaciens strain, LBA4404 is the most frequent, which has been used to introduce binary vectors [13]. In the last published protocol, EHA105 was applied more frequently [14- 16]. Plant expression vectors pCAMBIA2301 was used to identify candidate gene’s expression pattern, pBI121 was used to candidate gene function study, pK402 for gene editing [17-20]. Different explants have been used for transformation, such as leave [21-26]. internodes, tubers and petioles . Kanamycin is the most popular selective agent in published protocols. Herbicide resistance genes, phosphinothricin (bar) and glyphosate [N- (phosphonomethyl) glycine] (epsps) worked as selectable markers and introduce herbicide resistance traits into the transgenic plants as valuable traits.
The present study presents a streamlined and effective potato transgenic protocol utilizing A. tumefaciens for the Desiree cultivar. Drawing upon existing published methodologies, redundant steps and processes were eliminated to enhance efficiency. Various explants from the Desiree cultivar were evaluated, and the protocol was further assessed with two diploid materials, employing different types of explants, vectors, and selective agents. Positive transgenic plants from tetraploid Desiree and two diploid materials were confirmed via GUS histochemical assay and PCR analysis. Optimization of multiple variables has established a straightforward and efficient potato transgenic system tailored for Desiree [21-39].
Materials and Methods
Mediums Components Information
The components of potato growth media used in this research are listed in Table 1.
|
Reagents |
Brand |
Product ID |
Mediums |
|||||||
|
|
|
|
LB |
YEP |
INF |
COA |
SE |
SEG |
MS30T |
MS30 |
|
Yeast Extract |
OXOID |
LP0021 |
5g/L |
10g/L |
- |
- |
- |
- |
- |
- |
|
Tryptone |
OXOID |
LP0042 |
10g/L |
- |
- |
- |
- |
- |
- |
- |
|
NaCl |
Solarbio |
S8210 |
10g/L |
5g/L |
- |
- |
- |
- |
- |
- |
|
Bacteriological Peptone |
OXOID |
LP0137 |
- |
10g/L |
- |
- |
- |
- |
- |
- |
|
Agar |
Sigma-Aldrich |
A1296 |
15g/L |
15g/L |
9g/L |
9g/L |
9g/L |
9g/L |
7g/L |
7g/L |
|
Murashige & Skoog Basal Medium with Vitamins |
PhytoTech Labs |
M519 |
- |
- |
4.43g/L |
4.43g/L |
4.43g/L |
4.43g/L |
4.43g/L |
4.43g/L |
|
Sucrose |
SINOPHARM |
10021418 |
- |
- |
20g/L |
20g/L |
20g/L |
20g/L |
30g/L |
30g/L |
|
Acetosyringone |
Solarbio |
A8110 |
- |
- |
80mg/L |
80mg/L |
- |
- |
- |
- |
|
trans-Zeatin-riboside |
Solarbio |
Z8040 |
- |
- |
- |
1mg/L |
2mg/L |
2mg/L |
- |
- |
|
1-Naphthaleneacetic acid |
Sigma-Aldrich |
N0640 |
- |
- |
- |
2mg/L |
0.01mg/L |
0.02mg/L |
- |
- |
|
Timentin |
Solarbio |
T8660 |
- |
- |
- |
- |
200mg/L |
200mg/L |
100mg/L |
- |
|
Kanamycin |
Solarbio |
K8020 |
- |
- |
- |
- |
50mg/L |
100mg/L |
100mg/L |
- |
|
Gibberellic Acid (GA3) |
Solarbio |
G8040 |
- |
- |
- |
- |
- |
0.02mg/L |
- |
- |
|
pH |
|
|
|
|
|
|
|
|
|
|
|
(Adjusted by KOH) |
|
|
7.0 |
7.0 |
5.8 |
5.8 |
5.8 |
5.8 |
5.8 |
5.8 |
Table 1: Medium components used at different stages of genetic transformation
Plant Materials and Growth Condition.
The plantlets of the tetraploid variety Solanum tuberosum L. cv. Desiree, and two diploid lines, CIP703541 and CIP705079, were maintained on MS30 growth medium. The growth conditions were as follows: a light period of 16 hours with an intensity of approximately 16000 lux, followed by 8 hours of darkness. The temperature during light exposure was 21°C, while it was 20°C during the dark period. Details regarding the LED light spectrum can be found in Supplementary Figure S1b. All potato materials (Desiree, CIP703541, CIP705079) utilized in this study were sourced from the Breeding Resources Center (BRC) at the CIP China Center for Asia-Pacific, Beijing, China.
A. Tumefaciens Strains and Vectors
A. tumefaciens strain EHA105 carried vector pCAMBIA2301, pBI121, CBE and PE, respectively, which were stored in glycerol at -80°C. All vectors are binary vectors for plant transformation, pCAMBIA2301 and pBI121 containing GUS and NPT-II genes, and CBE and PE containing CAS9 and NPT-II genes. The maps of vectors are shown in Supplementary Figure S2.
A. Tumefaciens Mediated Genetic Transformation
After long-term storage, the A. tumefaciens was revived in a shaker at 28 °C, 150 rpm for 1 day in a YEP lipid medium. About 10 mL overnight incubated A. tumefaciens was centrifuged at 3500 rpm in a 50 ml tube for 10 min, the supernatant was discarded, and the pellet was re-suspended in about 30 mL volume INF medium containing 100mg/L Acetosyringone, which made the A. tumefaciens suspension concentration about 0.5, measured using a spectrophotometer adjusted to measure at OD550. About 30 explants, such as internodes, petioles, or leaf discs, were cut from 3-4 weeks-old healthy plants. The internodes contained no buds, and leaf samples were cut into discs. All explants were soaked in an A. tumefaciens suspension for 20 min with gentle shaking. The soaked explants were transferred to a COA medium using tweezers. Explants were placed on the surface of the COA medium. Residual bacterial suspension on the surface of explants, was removed using a pipette and air drying in an ultra-clean bench for 30 minutes. The plates were sealed by parafilm, and cultures were maintained in the dark at 21 °C for two days. The explants were then transferred to SE or SEG medium with kanamycin or chlorsulfuron under weak light (Light parameter shown in Supplementary Figure S1a). These were left to grow in the incubator under the same growth condition described in the 1.2 section. The explants were transferred to a fresh medium every two weeks to maintain selection pressure. After about four weeks, the shoots regenerated at the edges of the explant’s callus. The large shoots, about 2 cm in length, were dissected and transferred to MS30T medium for rooting.
GUS Histochemical Assay
Leaf samples (about 4 mm2) from seedlings were dipped in the GUS staining solution Kit (Biosharp, BL622A). All samples were incubated in stained solution at 37 °C overnight. Then, after 5 times applications of 80% acetone at room temperature, the leaf’s green coloration would fade. Finally, samples were stored in 75% ethanol for processing [40].
DNA Amplification Identified The Positive Transgenic Plantlets.
The leaf samples of regeneration seedlings, which were produced by transformation of PE and CBE, were collected in 2 ml Eppendorf tubes under lipid nitrogen and ground into powder. The genomic DNA was extracted using the CTAB method following Porebski’s protocol with some modifications [41]. The regenerated seedlings’ DNA, a positive control sample, and two negative control samples (double distilled water and receipt material) were identified by amplification marker gene NPT-II -relative special segment. The reaction program was shown as follows: denaturation at 94 °C for 4 min, followed by 35 cycles of denaturation at 94°C for 30, annealing at 55°C for 30 s, extension at 72°C for 30 s, a final extension at 72°C for 5 min. The primers’ sequence for PE transgenic seedlings is forward primer MMLV-06F: tgaggagggcctgcagcataactgc, reverse primer 26pR: gcctaatcattctaatcctgggaca; for the CBE vector, forward primer 35S: gacgcacaatcccactatcc, reverse primer NPTII-R: gatgtttcgcttggtggtcg.
Ploidy Analysis
Flow cytometric methods determined the ploidy level of regenerated plants. Crude samples of nuclei were prepared from the leaves of plantlets chopped in 1 ml phosphate-buffered saline (pH7.5) containing 0.5% Triton X-100 with DAPI and filtered by 75μm filter. The ploidy of cells was examined using a flow cytometer (Sysmex Shanghai Ltd. CyFlow Cube8). The isolated plant nuclear groups had characteristic fluorescence emission peaks. The DNA content of plantlets regenerated by diploid receptors was compared to potato material PL4 42, which worked as a diploid control, with the lowest peak corresponding to 2C nuclei and the remaining peaks representing 4C and 8C nuclei, respectively. Plantlets regenerated from Desiree were compared to their recipient, with the lowest peak corresponding to 4C nuclei. Noise signals generated by subcellular debris were eliminated by gating [42].
Results
Tetraploid Material Desiree Produced Higher Transgenic Efficiency and Lower Ploidy Change Ratio Compared 3o Diploid Materials.
|
Recipients |
Vectors |
Explants number |
Regeneration seedlings number |
Regeneration ratio |
Positive seedlings number |
Positive efficiency |
|
CIP703541 |
pCAMBIA2301, pBI121 |
80 |
53 |
66.25% |
3 |
3.75% |
|
CIP705079 |
pCAMBIA2301, pBI121 |
71 |
19 |
26.76% |
6 |
8.45% |
|
Desiree |
pCAMBIA2301, pBI121 |
154 |
103 |
66.88% |
37 |
24.03% |
Table 2: The Different Transgenic Efficiency Between Potato Receipts
For diploid material, CIP703541, 80 internodal and petiole sections (about 5 mm in length) were inoculated with EHA105 carrying the plant binary expression vector pCAMBIA2301 or pBI121. Of all the excised sections, about half regenerated with shoot growth. Then, following incubation in the GUS staining buffer, three shoots developed a blue coloration (transformation efficiency of 3.75%). The same transformation protocol was used for diploid CIP70507. Using 71 sections, 19 shoots regenerated, and 6 shoots acquired a blue color following GUS straining (8.45% efficiency). Finally, for the tetraploid cultivar Desiree, only using stem internodes, 154 sections were excised, producing 103 shoots. Following GUS staining, 29 shoots turned blue (24.03% efficiency) (Table 2). Despite the need to use stem internode and petiole internode sections for CIP703541 and CIP70507 transformations, which may influence the effectiveness of inoculation and shoot regeneration, efficiency results tend to indicate that Desiree's tetraploid material provides higher positive transgenic efficiency. The ploidy of regenerated shoots, including positive and negative transgenic materials, was analyzed by flow cytometry. Among a total of 91 samples, 71 were derived from Desiree, and the remaining 20 were from CIP705079 and CIP703541. For the diploid materials, 9 plants remained diploid after regeneration, giving a ploidy conversion rate of 55%. For Desiree, the ploidy conversion rate was 4% (Figure 1). Thus, the results indicate a greater rate of ploidy change, about half, in the diploids than in the tetraploid. The tetraploid material demonstrated greater ploidy stability after regeneration.

Figure 1:
The ploidy changed after regeneration for diploid materials CIP705079, CIP703541 and tetraploid material, Desiree.
(a) The DNA content of leaves’ nucleic acid from regeneration seedlings from diploid receipt CIP703541 by this transgenic system. Seedling
CIP703541-T193 is a diploid material, keep the same ploidy as receipt. Meanwhile, CIP703541-T76 is a tetraploid material. Diploid material PL4 worked as diploid control.
(b) The DNA content of leaves’ nucleic acid from regeneration seedlings from tetraploid receipt Desiree by this transgenic system. Seedling
Desiree-T96 is tetraploid, which is the stable regenerated seedling. Meanwhile, the Desiree-T80 is ploidy changed material. Desiree worked as tetraploid control.
The green arrow indicated 2C gate, red was for 4C gate, blue indicated undefinable peek.
SEG or SE Regeneration Medium Provided High Positive Transgenic Efficiency, Above 17%.
|
Medium |
Explants number |
Regeneration seedlings number |
Regeneration ratio |
Positive seedlings number |
Positive efficiency |
|
SE |
77 |
66 |
85.71% |
19 |
24.68% |
|
SEG |
105 |
45 |
42.86% |
18 |
17.14% |
Table 3: The different transgenic efficiency between selection media
Compared to SEG medium, SE just have half of NAA concentration and lack of gibberellic acid. When SE medium was used for regeneration, a total of 77 Desiree explants were infected by A. tumefaciens. Of these, 66 shoots regenerated, and 19 leaf samples turned blue after staining with GUS stain buffer, resulting in an efficiency of 24.68%. In another protocol using SEG medium, 105 explants were infected by A. tumefaciens, 45 shoots were regenerated. After GUS testing, 18 shoots appeared blue giving an efficiency of 17.14%. These preliminary results showed that when SE or SEG medium were used as single-regeneration media, both achieved high transformation efficiency rates exceeding 17%, which is comparable to a similar published protocol [43]. Nevertheless, overall, the performance of the SE medium was better than that of SEG (see Table 3).
Explants Derived from Leaves are More Suitable Than Internodes for the Desiree Transgenic System
It is easier for leaf discs to regenerate shoots than internodes. Shoots regenerated from leaf discs 28 days after transfer to SEG medium (Figure 2a1, 2b1). However, only calli appeared on the internodal sections. After 45 days, nearly 85% of leaf discs generated vigorous shoots (Figure 2a2). At the same time, for the internodes, almost 50% developed calli (Figure 2b2). After evaluating positive efficiency through GUS protein analysis, the results showed that, for the Desiree potato variety, leaf discs were more suitable as explants for transgenesis using SEG medium with A. tumefaciens. In total, 9 separate transgenic operations using PE and CBE for leaf discs were conducted, with 760 leaf discs used as explants. The regenerative rate ranged from 6.25% to 200%, averaging 53.14%. Meanwhile, 72 internodes were used as explants in 3 different operations, with a regenerative rate ranging from 0% to 79.41% (Figure 2c). These findings suggest that, compared to internodes, leaf discs are more stable as explants.

Figure 2:
The different of shoot regeneration between Desiree’s leaf discs and internodes explants
(a1-2) The regeneration of leaf discs explants. (a1) Leaf discs transfer to SEG medium at the first day. (a2) About 75% leaf discs explants regenerated shoots at 45 days after transferred to SEG medium.
(b1-2) The regeneration of internodal explants. (b1) Internodal explants transfer to SEG medium at the first day. (b2) About 30% internodal explants appeared shoots regeneration at 45 days after transfer to SEG medium. The red allows indicated internodal explants.
(c) The different regeneration ratio between leaf discs and internodal explants on separated experiments.
20 μg/L Chlorsulfuron is More Suitable for Selection than 40 μg/L.
To find the suitable selection concentration for chlorsulfuron, 294 Desiree explants were transferred to SEG medium containing 20 μg/L chlorsulfuron (without kanamycin) for callus induction. After 20 days of culture, by visual inspection, 136 remained healthy (Figure 3a) with a survival rate of 52.5%. Concurrently, 67 Desiree explants were transferred to SEG with 40 μg/L chlorsulfuron and without kanamycin; 20 days later, nearly all explants turned black and died (Figure 3a). Giving a survival rate is 3.36% (Figure 3b, Supplementary figure S3). Therefore, it is possible to deduce that when chlorsulfuron is used as a selection agent, 20 μg/L in the SEG medium is more suitable than 40 μg/L for Desiree.
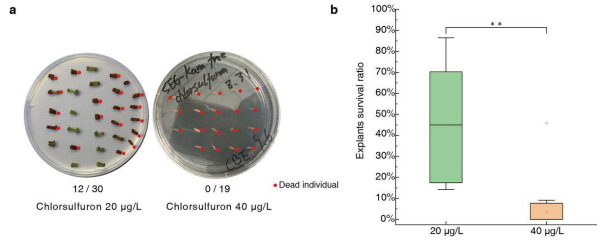
Figure 3:
The different survival ratio of explants under SEG medium with 20 μg/L and 40 μg/L chlorsulfuron respectively.
(a) The left plate shown explants on SEG plus 20 μg/L chlorsulfuron (without kanamycin) after 20 days culture. The right plant shown explants on SEG plus 20 μg/L chlorsulfuron (without kanamycin) after 20 days culture. Black explants indicated dead individual, marked by red dot.
(b) The different survival ratio of explants under SEG medium with 20 μg/L and 40 μg/L chlorsulfuron under separation trails.
The Transgenic System is Stable and Highly Efficient for Desiree Using Different Vectors.
In this study, four different binary vectors were assessed within the experimental system. The regeneration rates varied significantly among the vectors, with the CBE vector displaying the lowest rate at 31.22%, while the highest rate of 83.33% was observed when pBI121 was used as the vector for introduction. Selected seedlings underwent screening via GUS staining or PCR analysis to identify positive plantlets (Figure 4a, 4b). The results revealed an average positive rate of 72.36%. Notably, the lowest ratio was associated with pBI121 (50%), whereas the highest ratio was achieved with PE, reaching 96.00%. These findings suggest that the choice of backbone vector influences the shoot regeneration ratio. Despite variations among vectors, all regeneration rates exceeded 30%, with an average positive transgenic ratio exceeding 70%. This surpasses the efficiencies reported in common protocols, indicating the stability and high positive transgenic efficiency of this experimental system [43,44].

Figure 4:
The transgenic efficiency of Desiree work as receipt and infected by different vectors via A. tumefaciens
(a) The GUS staining assay for regeneration seedlings’ leaf. Leaf samples appeared blue were identified as positive transgenic line.
(b) PCR method identified the transgenic seedlings, which were regenerated from explants infected by A. tumefaciens with PE vector. ddH2O and Desiree worked as negative control, P0967 worked as positive control, sample 5 is a positive seedling.
(c) PCR method identified the transgenic seedlings, which were regenerated from explants infected by A. tumefaciens with CBE vector. ddH2O and Desiree samples worked as negative control, P0967 worked as positive control, sample 1, 2, 3 is positive seedlings. Sample 4 is a negative seedling.
The Potato Transgenic System Via A. Tumefaciens for Desiree is Simple and Time-Saving.
By eliminating the explant preculture process and employing a combination of easily regenerated explants, namely leaf discs, this system (Figure 6) demonstrated a rapid shoot emergence of approximately 0.5 cm in length within about 31 days for Desiree when inoculated with A. tumefaciens. This timeframe is notably quicker than certain traditional methods [45,46]. The preparation of A. tumefaciens with the target vector and explants required a single day, followed by a two-day co-culture on COA medium. Subsequently, it took 28 days for the first shoots, approximately 0.5 cm in length, to emerge on the callus maintained on the SEG medium. Thus, the total duration for shoot regeneration was 31 days. Roots typically emerged 10 days after transfer to the rooting medium, resulting in a total process completion time of 58 days. In summary, this system, through the elimination of the preculture process, utilization of a single-regeneration medium, and the use of leaf discs as suitable explants, streamlines operations and saves time.

Figure 5
Transgenic brief pipeline
Discussion
The Agrobacterium-mediated transgenesis method facilitates the creation of new materials transformed with desirable functional genes. This method serves as a robust tool for gene functional analysis, enabling the generation of genetically modified materials crucial for supporting basic research. Moreover, it holds immense potential for creating new varieties with modified gene functions to improve traits. Examples include drought tolerance, rapid tuber bulking for increased yield, and enhancement of various tuber qualities. Thus, the transgenic method utilizing A. tumefaciens stands as a pivotal technology for molecular research and breeding endeavors. Globally, numerous research groups are closely focused on successfully establishing potato transgenic systems based on A. tumefaciens, highlighting its significance in the scientific and agricultural communities.
In this study, the transgenic system proved highly efficient and time-saving for Desiree. However, its efficacy was notably lower for diploid materials. Significantly, we omitted the pre-culture procedures typically employed in some protocols (references 18 and 20), which typically require about one week for implementation. Instead, in this methodology, the explants were directly inoculated with A. tumefaciens. Following a two-day co-cultivation period, the explants were transferred to selection and regeneration medium. As a result, this protocol offers simplicity, with positive transgenic plants being produced approximately four weeks after inoculation.
Numerous factors influence potato transgenic efficiency via A. tumefaciens, with four key elements being particularly significant: the A. tumefaciens strain, potato recipients, culture medium, and growing environment.
We utilized A. tumefaciens strain EHA105 containing plant binary expression vectors for inoculation. The transgenic efficiency for Desiree’s disc eaves reached 68.80%, demonstrating that EHA105 efficiently introduces T-DNA into Desiree.
The vitality of A. tumefaciens is influenced by various factors, in-cluding culture medium, growth period, and environmental condi-tions. In our study, we employed YEP medium for A. tumefaciens culture, known for its suitability. Following overnight recovery, EHA105 exhibited robust vitality with rapid multiplication rates. Bacterial suspensions were then prepared in INF liquid medium, with an OD550 of approximately 0.5 optimized to ensure sufficient A. tumefaciens for explant inoculation while minimizing antibiotic effects during subsequent stages.
Recipient genotype significantly impacts potato transgenic efficiency. Desiree, frequently utilized as a recipient, demonstrated transformation efficiency of up to 68.80% in our study. However, diploid materials exhibited lower transformation efficiency, indicating a need for more suitable culture media. While our protocol utilizes a one-step regeneration medium (SEG medium), previous studies employing a two-step regeneration media protocol reported higher efficiencies for diploid materials.
Leaf discs and internodes were used to evaluate positive transgene efficiency in our study. Desiree's leaf discs exhibited higher efficiency than internodes, potentially due to differences in phytohormone distribution. Both internodes and petioles were successfully transformed, highlighting their suitability for transformation. Explants were sourced from 3–4-week-old plants, as older plants exhibit reduced regeneration ability.
The composition of culture media, including inoculation, co-cultivation, callus-induction, and shoot-induce media, significantly impacts transformation efficiency. Callus-induction medium is particularly crucial, containing antibiotics for selection and phytohormones to induce callus and shoot formation. In our study, SEG medium supplemented with NAA as an auxin and zeatin as a cytokinin was employed, providing optimal conditions for regeneration.
Our study employed a direct regeneration-single-stage technique for Desiree, simplifying the process and enhancing efficiency. However, for diploid materials, a two-stage technique may be more suitable, removing the auxin component to induce shoot formation.
While regulatory authorities have addressed NPT-II biosafety concerns, alternative selectable markers such as hygromycin may be considered. Chlorsulfuron was used as a selective agent in our study, with a concentration of 20μg/L proving suitable for selection.
As demonstrated here, eliminating unnecessary processes such as explant preculture streamlines the transformation process. Following inoculation, reducing A. tumefaciens concentration minimizes contamination risks during subsequent steps. Our simplified approach effectively decreased contamination risks and enhanced efficiency.
Somaclonal variations may occur during in vitro cultures, potentially impacting transgenic plants. Diploid materials in our study exhibited a high ploidy change ratio, emphasizing the need for careful selection.
Our study advances a simple and efficient potato transformation system via A. tumefaciens for Desiree, supporting gene functional research and biotechnology breeding endeavors. However, it underscores the importance of considering recipient genotypes and optimizing culture media for different potato materials.
Declarations
Author contributions
JL and CX prepared the manuscript and completed all trials. YL provided background information support with manuscript preparation. ME and PK proofread this manuscript for content and format and gave quite professional revision comments. QL, YJ and YF provided scientific suggestions. PY, ZZ, SC and YS support analyzed the data, provided key potato materials. JL and LW led the progress of this project, designed the experiments.
Acknowledgement
This work was supported by Scientic and Technological Innovation in Modern Seed Industry in Hebei Province (Grant No. 21326320D), the National Natural Science Foundation of China (Grant No, 32361143517), the China Agriculture Research System (CARS-09), the Hebei Potato Industry Collaborative Innovation Center, and Distinguished Professor of Hebei Northern University.
Conflict of Interest
Authors declare no conflict of interest.
Additional information
The data supporting the findings of this study are available in the article and supplementary materials. All data are available from the corresponding author.
Statement on Compliance of Plant Experiments
We solemnly declare that all work related to plant experiments and protocols has been conducted in accordance with the pertinent institutional, national, and international guidelines and legislation.
References
- Berchin, I. I., Silva de Amorim, W., & de Andrade Guerra,J. B. S. O. (2019). Food Security. In Sustainable Cities and Communities (pp. 1-9). Springer, Cham.
- Sharma, S., Kumar,A., Dhakte, P., Raturi, G., Vishwakarma, G., Barbadikar, K. M., ... & Deshmukh, R. (2023). Speed breeding opportunities and challenges for crop improvement. Journal of plant growth regulation, 42(1), 46-59.
- Shewry, P. R., Jones, H. D., & Halford, N. G. (2008). Plant biotechnology: transgenic crops. Food Biotechnology, 149-186.
- Fehér, A., Felföldi, K., Preiszner, J., & Dudits, D. (1991). PEG-mediated transformation of leaf protoplasts of Solanum tuberosum L. cultivars. Plant Cell, Tissue and Organ Culture, 27(1), 105-114.
- Romano, P., Fiore, C., Paraggio, M., Caruso, M., & Capece,A. (2003). Function of yeast species and strains in wine flavour. International journal of food microbiology, 86(1-2), 169-180.
- Ooms, G., Karp, A., & Roberts, J. (1983). From tumour to tuber; tumour cell characteristics and chromosome numbers of crown gall-derived tetraploid potato plants (Solanum tuberosum cv.‘Maris bard’). Theoretical and Applied Genetics, 66(2), 169-172.
- Perlak, F. J., Stone, T. B., Muskopf, Y. M., Petersen, L. J.,Parker, G. B., McPherson, S. A., ... & Fischhoff, D. A. (1993). Genetically improved potatoes: protection from damage by Colorado potato beetles. Plant molecular biology, 22(2), 313-321.
- USDA-APHIS (2015) Petitions for determination of nonregulated status, edited,http://www.aphis.usda.gov/ biotechnology/petitions_table_pending.shtml.
- Kaniewski, W. K., & Thomas, P. E. (2004). The potato story. AgBioForum, 7(1&2), 41-46.
- Hofvander, P. (2004). Production of amylopectin and high-amylose starch in separate potato genotypes (No. 495).
- Meier, A., & Halterman, D. A. (2016). Structural variation within the potato Ve gene locus and correlation with molecular marker analysis. Crop Science, 56(6), 3133-3142.
- Kaur, R. P., & Devi, S. (2019). In planta transformation in plants: a review. Agricultural Reviews, 40(3), 159-174.
- Vinterhalter, D., Zdravkovic-Korac, S., Mitic, N., Dragicevic, I., Cingel, A., Raspor, M., & Ninkovic, S. (2008). Protocols for Agrobacterium-mediated transformation of potato. Fruit, Vegetable and Cereal Science and Biotechnology, Global Science Books, 1-15..
- Elnahal, A. S., Li, J., Wang, X., Zhou, C., Wen, G., Wang, J., ... & Shan, W. (2020). Identification of natural resistance mediated by recognition of Phytophthora infestans effector gene Avr3aEM in potato. Frontiers in plant science, 11, 919.
- Ho, P. T., Byun, H. S., Vo, T. T., Lal, A., Kil, E. J., & Lee, S.(2022). First agroinoculation system development of Sweet potato symptomless virus on Nicotiana benthamiana.
- Ho, P. T., Byun, H. S., Vo, T. T., Lal, A., Lee, S., & Kil, E. J.(2023). Construction of an agroinfectious clone of a korean isolate of sweet potato symptomless virus 1 and comparison of its infectivity according to Agrobacterium tumefaciens strains in Nicotiana benthamiana. The plant pathology journal, 39(3), 255.
- Rahamkulov, I., & Bakhsh, A. (2020). Tissue-specific and stress-inducible promoters establish their suitability for containment of foreign gene (s) expression in transgenic potatoes. 3 Biotech, 10(10), 426.
- Kaur, A., Guleria, S., Reddy, M. S., & Kumar, A. (2020). A robust genetic transformation protocol to obtain transgenic shoots of Solanum tuberosum L. cultivar ‘Kufri Chipsona 1’. Physiology and Molecular Biology of Plants, 26(2), 367-377.
- Wang, K., Zhang, N., Fu, X., Zhang, H., Liu, S., Pu, X., ... & Si, H. (2022). StTCP15 regulates potato tuber sprouting by modulating the dynamic balance between abscisic acid and gibberellic acid. Frontiers in Plant Science, 13, 1009552.
- Ye, M., Peng, Z., Tang, D., Yang, Z., Li, D., Xu, Y., ... &Huang, S. (2018). Generation of self-compatible diploid potato by knockout of S-RNase. Nature Plants, 4(9), 651-654.
- Amiri, A. N., & Bakhsh, A. (2019). An effective pest management approach in potato to combat insect pests and herbicide. 3 Biotech, 9(1), 16.
- Bakhsh, A., Hussain, T., Rahamkulov, I., Demirel, U., & Çaliskan, M. E. (2020).Transgenic potato lines expressing CP4-EPSP synthase exhibit resistance against glyphosate. Plant Cell, Tissue and Organ Culture (PCTOC), 140(1), 23-34.
- De Block, M. (1988). Genotype-independent leaf disc transformation of potato (Solanum tuberosum) using Agrobacterium tumefaciens. Theoretical and applied genetics, 76(5), 767-774.
- Gustafson, V., Mallubhotla, S., MacDonnell, J., Sanyal-Bagchi, M., Chakravarty, B., Wang-Pruski, G., ... & Regan,S. (2006). Transformation and plant regeneration from leaf explants of Solanum tuberosum L. cv.‘Shepody’. Plant Cell, Tissue and Organ Culture, 85(3), 361-366.
- Rocha-Sosa, M., Sonnewald, U., Frommer, W., Stratmann, M., Schell, J., & Willmitzer, L. (1989). Both developmental and metabolic signals activate the promoter of a class I patatin gene. The EMBO journal, 8(1), 23-29.
- Shahin, E. A., & Simpson, R. B. (1986). Gene transfer system for potato. HortScience, 21(5), 1199-1201.
- Beaujean, A., Sangwan, R. S., Lecardonnel, A., & Sangwan-Norreel, B. S. (1998). Agrobacterium-mediated transformation of three economically important potato cultivars using sliced internodal explants: an efficient protocol of transformation. Journal of experimental Botany, 49(326), 1589-1595.
- Ducreux, L. J., Morris, W. L., Hedley, P. E., Shepherd, T., Davies, H. V., Millam, S., & Taylor, M. A. (2005). Metabolic engineering of high carotenoid potato tubers containing enhanced levels of β-carotene and lutein. Journal of experimental Botany, 56(409), 81-89.
- Heeres, P., Schippers-Rozenboom, M., Jacobsen, E., & Visser,R. G. (2002). Transformation of a large number of potato varieties: genotype-dependent variation in efficiency and somaclonal variability. Euphytica, 124(1), 13-22.
- Newell, C. A., Rozman, R., Hinchee, M. A., Lawson, E. C., Haley, L., Sanders, P., ... & Fraley, R. T. (1991). Agrobacterium-mediated transformation of Solanum tuberosum L. cv.‘Russet Burbank’. Plant Cell Reports, 10(1), 30-34.
- Orbegozo, J., Roman, M. L., Rivera, C., Gamboa, S., Tovar,J. C., Forbes, G. A., ... & Ghislain, M. (2016). Rpi-blb2 gene from Solanum bulbocastanum confers extreme resistance to late blight disease in potato. Plant Cell, Tissue and Organ Culture (PCTOC), 125(2), 269-281.
- Roman, M. L., Izarra, M., Lindqvist-Kreuze, H., Rivera, C., Gamboa, S., Tovar, J. C., ... & Ghislain, M. (2017). R/Avr gene expression study of Rpi-vnt1. 1 transgenic potato resistant to the Phytophthora infestans clonal lineage EC-1. Plant Cell, Tissue and Organ Culture (PCTOC), 131(2), 259-268.
- Kumar, A., Miller, M., Whitty, P., Lyon, J., & Davie, P. (1995). Agrobacterium-mediated transformation of five wild Solanum species using in vitro microtubers. Plant cell reports, 14(5), 324-328.
- Sheerman, S., & Bevan, M. W. (1988). A rapid transformation method for Solanum tuberosum using binary Agrobacterium tumefaciens vectors. Plant Cell Reports, 7(1), 13-16.
- Stiekema, W. J., Heidekamp, F., Louwerse, J. D., Verhoeven, H. A., & Dijkhuis, P. (1988). Introduction of foreign genes into potato cultivars Bintje and Desiree using an Agrobacterium tumefaciens binary vector. Plant Cell Reports, 7(1), 47-50.
- An, G., Watson, B. D., & Chiang, C. C. (1986). Transformation of tobacco, tomato, potato, and Arabidopsis thaliana using a binary Ti vector system. Plant Physiology, 81(1), 301-305.
- Kieu, N. P., Lenman, M., Wang, E. S., Petersen, B. L., & Andreasson, E. (2021). Mutations introduced in susceptibility genes through CRISPR/Cas9 genome editing confer increased late blight resistance in potatoes. Scientific Reports, 11(1), 4487.
- Tavazza, R., Ordas, R. J., Tavazza, M., Ancora, G., & Benvenuto, E. (1988). Genetic transformation of Nicotiana clevelandii using a Ti plasmid derived vector. Journal of plant physiology, 133(5), 640-644.
- Trujillo, C., Rodríguez-Arango, E., Jaramillo, S., Hoyos, R., Orduz, S., & Arango, R. (2001). One-step transformation of two Andean potato cultivars (Solanum tuberosum L. subsp. andigena). Plant Cell Reports, 20(7), 637-641.
- Lee, J. H., & Schöffl, F. (1997). GUS activity staining—a powerful tool in plant molecular biology. In Plant Molecular Biology Manual (pp. 23-32). Dordrecht: Springer Netherlands.
- Porebski, S., Bailey, L. G., & Baum, B. R. (1997). Modification of a CTAB DNA extraction protocol for plants containing high polysaccharide and polyphenol components. Plant molecular biology reporter, 15(1), 8-15.
- Galbraith, D. W., Harkins, K. R., & Knapp, S. (1991). Systemic endopolyploidy in Arabidopsis thaliana. Plant physiology, 96(3), 985-989.
- Bakhsh, A. (2020). Development of efficient, reproducible and stable Agrobacterium-mediated genetic transformation of five potato cultivars. Food Technology and Biotechnology, 58(1), 57-63.
- Ghislain, M., Byarugaba, A. A., Magembe, E., Njoroge, A., Rivera, C., Román, M. L., ... & Kiggundu, A. (2019). Stacking three late blight resistance genes from wild species directly into African highland potato varieties confers complete field resistance to local blight races. Plant Biotechnology Journal, 17(6), 1119-1129.
- Craze, M., Bates, R., Bowden, S., & Wallington, E. J. (2018). Highly efficient agrobacteriumâ?ÂÂmediated transformation of potato (Solanum tuberosum) and production of transgenic microtubers. Current protocols in plant biology, 3(1), 33-41..
- Trujillo, C., Rodríguez-Arango, E., Jaramillo, S., Hoyos, R., Orduz, S., & Arango, R. (2001). One-step transformation of two Andean potato cultivars (Solanum tuberosum L. subsp. andigena). Plant Cell Reports, 20(7), 637-641.