
Plant Biology Soil Health Journal(PBSHJ)
ISSN: 2996-2609 | DOI: 10.33140/PBSHJ


Short Communication - (2023) Volume 1, Issue 1
The Revision of Schizosaccharomycetaceae
2Department of Microbiology, Faculty of Biology and Biotechnology, University of Science, Vietnam National University-HCM City, 227 Nguyen Van Cu Street, Ward 4, District 5, Hochiminh City, Vietnam
3Microbial Diversity and Utilization Team, Thailand Bioresource Research Center (TBRC), National Center for Genetic Engineering and Biotechnology (BIOTEC), National Science and Technology Development Agency (NSTDA), 113 Thailand Science Park, Phaholyothin Road, Klong 1, Klong Luang, Pathumthani 12120, Vietnam
4Department of Biochemistry and Microbiology, Faculty of Pharmaceutical Sciences, Chulalongkorn University, 254 Phayathai Road, Wangmai, Pathumwan, Bangkok 10330, Thailand
5Institute for Fermentation, Osaka, 2-17-85 Juso-honmachi, Yodogawa-ku, Osaka 532-8686, Japan
6JICA Senior Overseas Volunteer, Japan International Cooperation Agency, Shibuya-ku, Tokyo 151-8559, Japan
7Laboratory of Applied Microbiology (Professor Emeritus), Department of Agricultural, Chemistry, Faculty of Agriculture, Shizuoka University, 836 Ohya, Suruga-ku, Shizuoka 422-8529, Japan
Received Date: Dec 01, 2023 / Accepted Date: Dec 22, 2023 / Published Date: Dec 30, 2023
Copyright: ©©2023 Yuzo Yamada, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation: Yamada, Y., Malimas, T., Vu, H. T. L., Yukphan, P., Tanasupawat, S., et al. (2023). The Revision of Schizosaccharomycetaceae. Plant Biol Soil Health J, 1(1), 61-66.
Abstract
Although the genus Hasegawaea was introduced along with the recognition of the genus Octosporomyces in the classification of fission yeasts, the two additional genera were neither accepted nor recognized. However, the genus Schizosaccharomyces sensu Kurtzman et Robnett was taxonomically heterogeneous-natured and corresponded to a higher-ranked taxon, i.e., a monotypic family. Thus, the following three genera were confirmed in the family Schizosaccharomycetaceae. The genus Schizosaccharomyces sensu stricto was comprised of Schizosacchromyces pombe, the genus Octosporomyces was of the four species, Octosporomyces octosporus, Octosporomyces osmophilus, Octosporomyces lindneri and Octosporomyces cryophilus and the genus Hasegawaea was of Hasegawaea japonica. In conclusion, the precise classification of microorganisms will not be able to be expected in the generic designation without the presence of taxonomic homogeneous-natured taxa. The phylogenetic distances have to be absolutely considered. Namely, the longer the distances are, the more taxonomic heterogeneous-natures will be increased in the resulting genus.
Keywords
Schizosaccharomyces Sensu Stricto, Octosporomyces, Hasegawaea, Fission Yeasts
Supplementary Abstract
|
The family Schizosaccharomycetaceae Beierinck et Klöcker Genus
|
Species S. pombe Lindner (1893) O. octosporus (Beijerinck) Kudriav-zev O. osmophilus (Brysch-Herzberg et al.) Vu et al. comb. nov. O. lindneri (Brysch-Herzberg et al.) Malimas et al. comb. nov. O. cryophilus (Helston et al.) Vu et al. comb. nov. H. japonica Yamada et Banno (1987) |
|
The family Schizosaccharomycetaceae Beierinck et Klöcker Genus 1. Schizosaccharomyces Lindner (1893) sensu Kurtzman et al. (1991) |
Species S. pombe Lindner (1891) S. octosporus Beeijerinck (1894) S. japonicus Yukawa et Maki (1931) S. cryophilus Helston et al (2010) S. osmophilus Brysch-Herzberg et al. (2019) S. lindneri Brysch-Herzberg et al. (2023) |
In the family Schizosaccharomycetaceae Beijerinck ex Klöcker, the genus Hasegawaea Yamada et Banno was introduced for Schizosaccharomyces japonicus, along with the recognition of the genus Octosporomyces Kudriavzev, based on the differences in ubiquinone isoprenologues as one of the most decisive criteria as well as in ascospore morphology [1-4]. The resulting three genera Schizosaccharomyces, Octosporomyces and Hasegawaea were thus taxonomically homogeneous-natured, i.e., morphologically, physiologically, chemotaxonomically, phylogenetically and genetically. However, the two generic names were neither accepted nor recognized taxonomically [5-7].
This paper describes the historical surveys in the classification of fission yeasts to retrieve the generic names of Hasegawaea and Octosporomyces from the phylogenetic and the phenotypic points of view.
• The family Schizosaccharomycetaceae Beijerinck ex Klöcker
• The type genus is Schizosaccharomyces. The three genera are included.
Genus I.
Schizosaccharomyces Lindner (MB 4905)
Four-spored, warty ascospores and ubiquinone-10 (Q-10)
Schizosaccharomyces pombe Lindner (1893) (MB 212377)
The type strain is NBRC 1628T (= CBS 356T = CLIB 833T).
KurtzmanandRobnett(1991)describedthatSchizosaccharomyces pombe and Schizosaccharomyces octosporus were more closely related to one another than were Saccharomyces kluyveri and Saccharomyces unisporus and that Schizosaccharomyces octosporus and Schizosaccharomyces japonicus were separated by a distance only 25% greater than found for outlying Saccharomyces species based on their phylogenetic tree. However, the two Saccharomyces species were later transferred to the separate two genera Lachancea and Kazachstania as Lachancea kluyveri and Kazachstania unispora (Kurtzman 2003), indicating that the Schizosaccharomyces species should also share the same fate. Therefore, the authors’ opinion rejecting the two generic names of fission yeasts completely lost its validity due to the mistakes in reference standard or reference control selection [8].
Yamada et al. (1993) determined the partial base sequences of fission yeasts in the three regions of 26S rRNA (positions 493-622, 130 bases, designated as region a) and positions 1611-1835, 225 bases, designated as region b) and 18S rRNA (positions 1451-1618, 168 bases, designated as region c) [9]. In the partial base sequencings, Schizosac-charomyces octosporus represented one base substitution, when compared with Schizosaccharomyces pombe, in region c. However, the maximum homologies were very low (75-77%) in region a and the base substitutions were very high (45-16) in region b, when compared with the remaining two species. In addition, Schizosacharo-myces japonicus represented very low maximum homologies and very high base substitutions in all the three regions, i.e., 75-84% maximum homologies in regions a, 45-37 base substitutions in region b, and 7-6 base substitutions in region c.
Actually, the phylogenetic branches among Schizosaccharomyces pombe, Octosporomyces octossporus and Hasegawaea japonica were much longer than that between Vanderwaltozyma polyspora (Kurtzman 2003) and Saccbaromyces cerevisiae used as reference standards (Fig. 1), indicating that the three species should be separated from one another at the generic level [8].
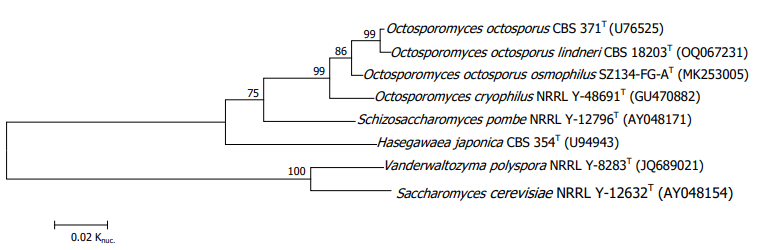
Figure 1: The phylogenetic tree of fission yeasts based on the 26S rRNA gene D1/D2 domain sequences for 543 bases. The evolutionary history was inferred using the neighbour-Joining method. Vanderwaltozyma polyspora and Saccharomyces cerevisiae were used as outgroup. The bootstrap test (1000 replicates) was shown next to the branches. For the construction of the phylogenetic tree, refer to Yamada et al.
Additionally, the calculated sequence similarities (84.4-90.4%) among the three species were lower than that (91.7%) (Vu et al. 2021; Yamada et al. 2022b) between Babjevia anomala (= Dipodascopsis anomala, Q-9) and Lipomyces starkeyi (Q-9) in the 26S rRNA gene D1/D2 domain sequences (Table 1) [10, 11]. By the way, the sequence simi-larity between Vanderwaltozyma polyspora (Q-6) and Saccharomyces cerevisiae (Q-6) was 93.8%.
|
Species |
1 |
2 |
3 |
4 |
5 |
6 |
|
1. Schizosaccharomyces pombe CBS 356T |
100 |
90.4 |
90.6 |
90.1 |
90.9 |
89.8 |
|
2. Octosporomyces octosporus CBS 371T |
|
100 |
98.1 |
99.4 |
95.2 |
84.4 |
|
3. Octosporomyces osmophilus SZ134-FG-AT |
|
|
100 |
97.9 |
96.4 |
85.0 |
|
4. Octosporomyces lindneri NRRK Y-4869T |
|
|
|
100 |
95.0 |
84.3 |
|
5. Octosporomyces cryophilus NRRL Y-48691T |
|
|
|
|
100 |
84.5 |
|
6. Hasegawaea japonica CBS 354T |
|
|
|
|
|
100 |
The percent sequence similarity was calculated for 589 bases. For the calculation, refer to Yamada (2022).
Table 1: The pair-wise 26S rRNA gene D1/D2 domain sequence similarity.
Thus, the genera Hasegawaea and Octosporomyces should be enough to be unequivocally accepted and retained in the classification of fission yeasts.
Genus II.
Octosporomyces Kudriavzev (MB 3551)
Basically eight-spored, smooth ascospores and ubiquinone-9 (Q-9)
The type species is Octosporomyces octosporus.
Four species are included.
1. Octosporomyces octosporus (Beijerinck) Kudriavzev (1960) (MB 335285)
Basionym: Schizosaccharomyces octosporus Beijerinck (1894).
The type strain is NBRC 10373T (= CBS 371T = CLIB 832T).
2. Octosporomyces osmophilus (Brysch-Herzberg, Tobias, Seidel, Wittmann, Fischer, Dlauchy et Péter) Vu, Yukphan, Tanasupawat, Mikata et Yamada comb. Nov.
Basionym: Schizosaccharomyces osmophilus Brysch-Herzberg, Tobias, Seidel, Wittmann, Fischer, Dlauchy et Péter, FEMS Yeast Res 19; foz038-10: 2019.
The type strain is CBS 15793T (= CLIB 3267T).
MycoBank number is 846278.
3. Octosporomyces lindneri (Brysch-Herzberg, Sipiczki, Jia, Seidel et Du) Malimas, Vu, Yukphan, Tanasupawat et Yamada comb. nov.
Basionym: Schizosaccharomyces lindneri Brysch-Herzberg, Sipiczki, Jia, Seidel et Du, Yeast Wiley 13: 2023.
The type strain is CBS 18203T.
MycoBank number is 849741.
4. Octosporomyces cryophilus (Helston, Box, Tang et Baumann) Vu, Yukphan, Tanasupawat, Mikata et Yamada comb. nov.
Basionym: Schizosaccharomyces cryophilus Helston, Box, Tang et Baumann, FEMS Yeast Res 10; 784: 2010.
The type strain is NRRL Y-48691T (= CBS 11777T = NBRC 106824T).
MycoBank number is 846279.
In the fission yeasts, the two new Schizosaccharomyces species were described [12, 13]. Of the two, Schizosaccharomyces osmophilus was phylogenetically very closely related to Octosporomyces octosporus (Fig. 1). The sequence similarity was 98.1% between the two species (Table 1), indicating that the new species was obviously classified in the genus Octosporomyces but not in the genus Schizosaccharomyces sensu stricto. The phylogenetic data obtained above was supported by Brysch-Herzberg et al. (2019) who calculated 1.8% base substitution (= 98.2% sequence similarity) [13]. However, the authors accommodated the new species to the genus Schizosaccharomyces sensu Kurtzman et Robnett, which is taxonomic-heterogeneous natured, but not to the genus Octosporomyces Kudriavzev, which is phylogenetically closely related and taxonomic-homogeneous natured. The calculated sequence similarity mentioned above was almost the same as that (98.0%) between Kockiozma suomiensis (=Zygozyma suomiensis, Q-8) and Myxozyma geophila (Q-8) [10, 11].
Recently, the new species, Schizosaccharomyces lindneri was described [14]. The species represented an extremely short phylogenetic branch to Octosporomyces octosporus (Fig. 1). The calculated sequence similarity was very high (99.4%), which was higher than that (98.1%) of Octosporomyces osmophilus (Table 1). Thus, it is reasonable that the species should be accommodated to the genus Octosporomyces.
Concerning Schizosaccharomyces cryophilus, the phylogenetic position of the species was not so closely related (Fig. 1). The sequence similarities were 95.2% and 96.4% respectively to Schizosaccharomyces octosporus and Schizosaccharomyces osmophilus (Table 1). The calculated sequence similarities were not so high, so that the species may be able to be accommodated to a different genus, as shown in the relationships between Babjevia anomala and Neoaidaea tothii, in which the calculated sequence similarity was 95.6% (Vu et al. 2021; Yamada et al. 2022b) and between Kloeckeraspora osmophila and Kloeckeraspora occidentalis, in which the calculated sequence similarity was 94.0% [15]. However, it is adequate at the present time to be temporarily classified in the genus Octosporomyces, since the species had ubiquinone-9 (Q-9) (Kaino et al. 2018) and an almost identical length of ITS1 (307 bp, internal transcribed spacer 1) to that (343 bp) of Schizosaccharomyces octosporus [16, 12]. Incidentally, Schizosaccharomyces pombe had 417 bp ITS1 and Schizosaccharomyces japonicus had 183 bp ITS1.
Genus III.
Hasegawaea Yamada et Banno (MB 25179)
Basically eight-spored, smooth ascospores without papillae, dimorphic growth, respiration
deficiency and no detectable ubiquinone (a trace amount of Q-10)
Hasegawaea japonica (Yukawa et Maki) Yamada et Banno (1987) (MB 132784)
Basionym: Schizosaccharomyces japonicus Yukawa et Maki (1931).
Synonym: Octosporomyces japonicus (Yukawa et Maki) Kudriavzev (1960).
The type strain is NBRC 1609T (= CBS 354 T = CLIB 3267T)
Schizosaccharomyces japonicus is especially distant phylogenetically, i.e., the species is the furthest removed from other species among the fission yeasts [17-19].
Liu et al. (2009) made phylogenomic analyses on the basis of the monophyly of Taphrinomycotina, including Schizosaccharomyces fission yeasts. In the phylogenetic tree, Schizosaccharomyces japonicus evolved first and separated from the remaining two species, Schizosaccharomyces pombe and Schizosaccharomyces octosporus, as described above [19]. However, there was a certain risk that a heteroge-neous-natured taxon was sometimes given taxonomically in the generic designation (Vu et al. 2021; Yamada et al. 2022b), when based on the monophyly only, and the resulting genus corresponded to a higher-ranked taxon, i.e., the monotypic family Schizosac-charomycetaceae, as discussed in the genus Wickerhamomyces Kurtzman et al. (2008), in which the genus Hansenula Sydow et Sydow (1919) was actually included [20].
In the monotypic family Schizosaccharomycetaceae, the genus Schizosaccharo-myces sensu actually had quite heterogeneous natures genetically, chemotaxonomically and physiologically [3, 21, 16, 7, 22, and 19]. In addition, it is of interest that the branch between Schizosaccharomyces japonicus and Schizosaccharomyces pombe was much longer than that between Saccharomyces (Q-6) and Pichia (Q-7) in the phylogenetic tree based on the concatenated sequences of 13 proteins (cox1, cox2, cox3, cob, atp6, atp9 and nad1, nad2, nad3, nad4, nad4L, nad5, nad6) encoded by mtDNA. The experimental data obtained above indicated that the genus Hasegawaea could not be rejected but retained.
A maximum-likelihood phylogeny of the fission yeasts from the concatenation-based analysis of 815 BUSCO amino acid genomic sequences (total 56, 2376 sites) was studied by Rhind et al. (2011). The resulting phylogenetic tree was very similar in topology to that of Liu et al. (2009) as well as to that mentioned above based on the 26S rRNA gene D1/D2 domain sequences (Fig. 1).
In the comparative study on wtf genes in the family Schizosaccharomycetaceae by De Carvalho (2020), it was emphasized that the only Schizosccharomyces japonicus did not have such genes [23]. And the evolutionary analysis indicated that a very similar topology to those described above was found in the resulting phylogenetic tree. Schizosaccharomyces japonicus was first distantly removed from the others, and then Schizosaccharomyces pombe was. In contrast, the branches were quite short among Schizosaccharomyces octosporus, Schizosaccharomyces osmophilus, Schizosaccharo-myces lindneri and Schizosaccharomyces cryophilus, indicating that the existence of three genera was confirmed and the latter four species were adequate to be accom-modated to the genus Octosporomyces [24].
Kurtzman (2003) introduced ‘clade’ or ‘phylogenetically circumscribed genus’ as generic concept. However, it was not yet perfect to get the taxonomic homogeneous-natured taxon for the genus. Namely, the branch lengths should be additionally considered in the phylogenetic trees.
In the seven ranks used for the systematics of plants and animals, the taxonomically homogeneous taxa can be available only when the lower-ranked two taxa, i.e., species and genus are used. Therefore, the present authors naturally selected a taxonomic homogeneous-natured taxon in the generic designation, since the longer the phylo-genetic distances are the more taxonomic heterogeneous-natures will be increased.
Thus, the genus Schizosaccharomyces Lindner sensu Kurtzman et Robnett (1991) corresponded exactly to the monotypic family that actually includes the phylogenetic-distant and the phenotypic-distinct species, as far as the genus is not divided into three, i.e., Schsizosaccharomyces Lindner (1893) sensu stricto, Octosporomyces Kudriavzev (1960) and Hasegawaea Yamada et Banno (1987).
Acknowledgements
The authors express their sincere thanks for citing a number of articles for detailed discussions.
Funding information
The authors received no specific grant from any funding agency.
References
- YAMADA, Y., & BANNO, I. (1987). Hasegawaea gen. nov., an ascosporogenous yeast genus for the organisms whose asexual reproduction is by fission and whose ascospores have smooth surfaces without papillae and which are characterized by the absence of coenzyme Q and by the presence of linoleic acid in cellular fatty acid composition. The Journal of General and Applied Microbiology, 33(3), 295-298.
- Yamada, Y., & Banno, I. (1989). A proposal to divide the genus Schizosaccharomyces lindner sensu yarrow into three separate genera. Yeast (Chichester), 5, 393-398.
- Yamada, Y., ARIMOTO, M., & KONDO, K. (1973).Coenzyme Q system in the classification of the ascosporogenous yeast genus Schizosaccharomyces and yeast-like genus Endomyces. The Journal of General and Applied Microbiology, 19(5), 353-358.
- MIKATA, K. (1987). Surface structure of ascospores of the genus Schizosaccharomyces. IFO Res. Commun., 13, 45-51.
- Kurtzman, C. P., & Robnett, C. J. (1991). Phylogenetic relationships among species of Saccharomyces, Schizosaccharomyces, Debaryomyces and Schwanniomyces determined from partial ribosomal RNA sequences. Yeast, 7(1), 61-72.
- Kurtzman, C. P., & Robnett, C. J. (1998). Identification and phylogeny of ascomycetous yeasts from analysis of nuclear large subunit (26S) ribosomal DNA partial sequences. Antonie van Leeuwenhoek, 73, 331-371.
- Vaughan-Martini, A., & Martini, A. (2011). Schizosaccharomyces Lindner (1893). In The yeasts (pp. 779-784). Elsevier.
- Kurtzman, C. P. (2003). Phylogenetic circumscription of Saccharomyces, Kluyveromyces and other members of the Saccharomycetaceae, and the proposal of the new genera Lachancea, Nakaseomyces, Naumovia, Vanderwaltozyma and Zygotorulaspora. FEMS yeast research, 4(3), 233-245.
- Yamada, Y. (1993). The phylogenetic relationships of fission yeasts based on the partial sequences of 18S and 26S ribosomal RNAs: the recognition of Hasegawaea Yamada et Banno along with Schizosaccharomyces Lindner. Bull. Fac. Agric. Shizuoka, 43, 29-38.
- Vu, H. T. L., Yukphan, P., Tanasupawat, S., & Yamada, Y. (2021). The revision of the family Lipomycetaceae. 27th Ann Meet JSMRS, 15.
- Yamada, Y., Vu, H. T. L., Yukphan, P., & Tanasupawat, S. (2022). The revision of Lipomycetaceae.
- Helston, R. M., Box, J. A., Tang, W., & Baumann, P. (2010). Schizosaccharomyces cryophilus sp. nov., a new species of fission yeast. FEMS yeast research, 10(6), 779-786.
- Brysch-Herzberg, M., Tobias, A., Seidel, M., Wittmann, R., Wohlmann, E., et al. (2019). Schizosaccharomyces osmophilus sp. nov., an osmophilic fission yeast occurring in bee bread of different solitary bee species. FEMS yeast research, 19(4), foz038.
- Brysch-Herzberg, M., Jia, G. S., Sipiczki, M., Seidel, M., Li, W., et al. (2023). Schizosaccharomyces lindneri sp. nov., a fission yeast occurring in honey. Yeast.
- Malimas, T., Vu, H. T. L., Yukphan, P., Tanasupawat, S., & Yamada, Y. (2023). The Reinstatement of the Genus Kloeckeraspora Niehaus (1932)(Apiculate Yeast).
- Kaino, T., Tonoko, K., Mochizuki, S., Takashima, Y., & Kawamukai, M. (2018). Schizosaccharomyces japonicus has low levels of CoQ10 synthesis, respiration deficiency, and efficient ethanol production. Bioscience, biotechnology, and biochemistry, 82(6), 1031-1042.
- Liu, Y., Leigh, J. W., Brinkmann, H., Cushion, M. T., Rodriguez-Ezpeleta, N., et al. (2009). Phylogenomic analyses support the monophyly of Taphrinomycotina, including Schizosaccharomyces fission yeasts. Molecular biology and evolution, 26(1), 27-34.
- Rhind, N., Chen, Z., Yassour, M., Thompson, D. A., Haas,B. J., et al. (2011). Comparative functional genomics of the fission yeasts. Science, 332(6032), 930-936.
- Aoki, K., Furuya, K., & Niki, H. (2017). Schizosaccharomyces japonicus: A Distinct Dimorphic Yeast among the Fission Yeasts. Cold Spring Harbor protocols, 2017(12), pdb-top082651.
- Malimas, T., Vu, H. T. L., Yukphan, P., Tanasupawat, S., & Yamada, Y. The genus Hansenula Sydow et Sydow (1919) nom. rev. and the genus Wickerhamomyces Kurtzman et al.(2008).
- Mikata, K. (1999). The ubiquinone system in Hasegawaea japonica (Yukawa et Maki) Yamada et Banno: A new method for identifying ubiquinone homologs from yeast cells. IFO Res. Comm., 19, 41-46.
- Sipiczki, M., Takeo, K., Yamaguchi, M., Yoshida, S., & Miklos, I. (1998). Environmentally controlled dimorphic cycle in a fission yeast. Microbiology, 144(5), 1319-1330.
- De Carvalho, M. (2020). wtf: killing gametes for for more than 110 million years.
- Malimas, T., Vu, H. T. L., Yukphan, P., Tanasupawat, S., Mikata, K., et al. (2022). The Revision of Schizosaccharomycetaceae.