


Research Article - (2026) Volume 4, Issue 1
Synergistic Effects of Silicon and Bio fertilizers on Growth and Water Stress Tolerance of Faba bean (Vicia faba L.)
Received Date: Feb 02, 2026 / Accepted Date: Mar 09, 2026 / Published Date: Mar 16, 2026
Copyright: ©2026 Elham. A. Badr, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation: Elkaramany. M. F., Saied. E. S., Abou Basha. D. M., Abdel-Kader. H. H., Elham. A. B. (2026). Synergistic Effects of Silicon and Bio fertilizers on Growth and Water Stress Tolerance of Faba bean (Vicia faba L.). J Water Res, 4(1), 01-07.
Abstract
Objective: This study aimed to evaluate the effectiveness of foliar application of silicon (Si) in improving growth, yield, and chemical composition of faba bean plants grown under sufficient and deficit irrigation conditions, with or without bio fertilizer application in sandy soils.
Methods: A field experiment was conducted at the National Research Centre experimental station, Nubaria District, Behira Governorate, Egypt, during two consecutive winter seasons (2023/2024). Silicon was applied as a foliar spray at rates of (0, 50, 75, and 100 ppm) under sufficient and deficit irrigation regimes, combined with bio fertilizer treatments. The experiment followed a split-block design arranged factorial. Growth traits, physiological parameters, yield components, and macro- and micronutrient contents were assessed.
Results: Silicon application significantly affected all studied parameters. The 75 ppm Si treatment achieved the highest chlorophyll content (SPAD), total water status (TWS %), and relative water content (RWC %). However, 100 ppm Si produced the greatest improvements in vegetative growth, seed yield, and nutrient accumulation (N, P, K, Ca, Fe, Zn, Mn, and Cu) in roots, shoots, and seeds. Sufficient irrigation significantly enhanced plant performance compared with deficit irrigation. The interaction between 100 ppm Si and sufficient irrigation resulted in maximum values for most traits, while the combined application of 100 ppm Si, sufficient irrigation, and bio fertilizer produced the highest fresh and dry biomass as well as seed protein content.
Conclusion: Foliar application of 100 ppm silicon integrated with adequate irrigation and bio fertilizer represents an effective strategy for improving faba bean productivity and nutritional quality under sandy soil conditions.
Keywords
Sufficient Irrigation, Deficit, Nutrient Content, Proline, Water Stress, Water use Efficiency, Chlorophyll Content (SPAD),Relative Water Content (RWC), Nutrient Accumulation, Seed Yield, Protein Content, Sustainable Agriculture.
Introduction
Faba bean (Vicia faba L.) is one of the most important leguminous crops cultivated in Egypt, It represents a staple component in human nutrition and is also widely used in animal feed, either as green pods or dry seeds [1]. In 2021, the cultivated area in Egypt reached 26,382 hectares, with an average seed yield of 3.257 tons per hectare [2]. Faba bean seeds are highly valued for their nutritional composition, containing 22–25% protein in dry seeds, in addition to carbohydrates, cellulose, minerals, and vitamin C [3]. Like other legume crops, faba beans contribute significantly to improved soil fertility through the fixation of more soil nitrogen (N) [4]. However, faba bean productivity in Egypt is severely constrained by water scarcity and the low fertility of sandy soils in newly reclaimed lands [5]. Water stress reduces plant water potential due to limited soil moisture, high evaporative demand, or resistance to water transport within plant tissues, leading to growth inhibition and stomatal closure .Improving water use efficiency under such conditions is therefore essential to sustain productivity under limited water resources. Silicon (Si), although not classified as an essential nutrient for most crops, has emerged as a promising element in enhancing plant tolerance to abiotic stresses [6]. Its beneficial effects are mainly attributed to deposition within plant tissues, improving structural integrity and stress resistance. Silicon enhances leaf water retention, maintains membrane stability, improves stomatal conductance, and strengthens xylem vessels under drought conditions [7]. Silicon can diminish the electrolyte leakage from plant leaves and therefore raised photosynthetic activity in plants grown under water stress conditions (Epstein 2011). Matoh et al., 2015 found that application of Si led to formation of a silica cuticle layer on epidermal tissue of leaf, which is responsible for greater leaf water potential under water deficit conditions. Different studies indicated the positive effect of Si application on growth, yield and quality of many plants, Lu et al., 2016; Sukkaew et al., 2016 and Gorecki and Danielski, 2018. Application of Si nanoparticles was used to improve flowering rates and yields in faba bean (Asmaa et al., 2024). Low water potential in the plant because of low soil water potential, high evaporative demand, and/or a significant barrier to water flow through the plant are the causes of water stress. The most sensitive processes in the crop, growth decrease, and stomatal closure are the two most affected processes by the water deficiency. However, there are other processes as well Victor et al. (2016). Egypt is known for using chemical fertilizers extensively due to its intensive farming practices.
As a result, it is often advised to apply biofertilizers concurrently, first to improve the biological, physical, and chemical properties of the soil and then to obtain high-quality agricultural products free of unwanted high concentrations of heavy metals and other pollutants (El kramany et al., 2007). The application of biofertilizer inoculants fostered nutrient-rich, high-yielding plants, underscoring the value of microbial inputs in sustainable agriculture. This integrated approach optimizes nitrogen fixation, supports plant growth, and improves soil fertility, positioning it as an effective strategy for increasing legume crop productivity (Fares, 2025). Therefore, the present study aims to evaluate the individual and combined effects of silicon and biofertilizers on growth attributes, physiological responses, yield components, and water use efficiency of faba bean plants grown under different irrigation regimes in sandy soil.
Materials and Methods
Experimental Design
Field experiment was carried out during the two successive winter seasons of 2022/2023 and 2023/2024 at the Experimental Station of National Research Centre, Nubaria District, Behira Governorate, Egypt. This experimental farm (latitude 30°30'1.4''N, and longitude 30°19'10.9''E, and mean altitude 21 m above sea level). The soil of both experiment sites (0 - 30 cm) were newly reclaimed sandy soils where physical and chemical properties of the soil determined according to Jackson is shown in Table (1) [8].
The experiment had 3 factors (silicon, irrigation and biofertilizer) contain 16 treatments in 3 replicates with a total of 48 plots. silicon located in main plots, irrigation in sub-plots with and without biofertilizer, combined analysis of two seasons was done. This work aimed to investigate the impact of spraying solution of silicon at different levels (0, 50, 75 and 100 ppm) of supplementation on growth, yield and their chemical composition of Faba bean variety (Nubaria-1), irrigation treatments were (sufficient 1300 m3/fed or deficit 600 m3/fed.) Crop evapotranspiration (ETc) was calculated according to the following formula: (ETc = Kc x ET0), the water requirements of Faba bean plant grown at Nubaria station are present in Table (2).
|
Soil characteristics |
1st season |
2nd season |
Soil characteristics |
1st season |
2nd season |
|
Particle size distribution (%) |
Available Macronutrients (mg kg soil-1) |
||||
|
Sand |
91.02 |
90.94 |
N (K- Chloride Ex.) |
11.26 |
11.32 |
|
Silt |
3.81 |
3.92 |
P (Na Bicarbonate Ex.) |
1.55 |
1.53 |
|
Clay |
4.73 |
4.61 |
K (Ammo. Acetate. Ex.) |
8.94 |
8.78 |
|
Texture class |
Sandy |
Sandy |
Soluble cations (meq/L) |
||
|
Na+ |
2.19 |
2.16 |
|||
|
pH (1:2.5) |
7.21 |
7.16 |
K+ |
1.06 |
1.12 |
|
Ca++ |
2.06 |
1.98 |
|||
|
EC (dS m-1) |
0.76 |
0.71 |
Mg++ |
1.71 |
1.64 |
|
Soluble anions (meq/L) |
|||||
|
Organic matter (%) |
0.37 |
0.35 |
CO3-- |
0.00 |
0.00 |
|
HCO3- |
2.76 |
2.65 |
|||
|
CaCO3 (%) |
0.91 |
0.89 |
Cl- |
1.17 |
1.23 |
|
SO4-- |
3.24 |
3.12 |
|||
Table 1: Some Physical and Chemical Properties of the Experimental Soil
|
Month |
Sept. |
Oct. |
Nov. |
Dec. |
Jun |
Total |
||||
|
Period |
15-30 |
|
1-30 |
|
1-30 |
|
1-31 |
|
1--15 |
|
|
ETo mm day 1 |
5.9 |
|
6.4 |
|
7 |
|
6.2 |
|
5.4 |
|
|
No. of day |
13 |
10 |
12 |
8 |
10 |
6 |
11 |
|
10 |
80 |
|
Kc |
|
0.53 |
|
0.88 |
|
1.09 |
|
0.72 |
|
|
|
Kr |
|
0.7 |
|
0.85 |
0.91 |
0.95 |
1.00 |
|
|
|
|
Etc/loc. mm day 1 |
1.86 |
2.02 |
4.07 |
4.45 |
5.90 |
5.46 |
3.79 |
3.30 |
|
|
|
Ks |
1.15 (87%) |
|
|
|
|
|
|
|
|
|
|
Eu |
1.11 (90%) |
|
|
|
|
|
|
|
|
|
|
Lr |
10% |
|
|
|
|
|
|
|
|
|
|
Lrg mm day 1 |
2.46 |
2.67 |
5.38 |
5.88 |
7.80 |
7.21 |
5.01 |
4.37 |
|
|
|
Lrg L Day-1 plant-1 |
0.77 |
0.83 |
1.681 |
1.838 |
2.437 |
2.254 |
1.567 |
1.366 |
|
|
|
Lrg L season-1 plant-1 |
10.98 |
5.10 |
27.18 |
17.03 |
46.91 |
25.52 |
27.54 |
18.84 |
|
120 |
|
Lrg L m3 season-1 plant-1 |
126.5 |
95.0 |
216.0 |
113.0 |
305.0 |
150.5 |
196.0 |
98.00 |
|
|
|
(I1) |
215.5 |
329.5 |
455.5 |
294 |
1300 m3 fed-1 |
|||||
Table 2: Water Requirement for Drip Irrigated Faba Bean Grown on Sandy Soil at Nubaria Behira, Governorate
Fertilizers application:
Bio fertilizers
Before transplanting, Faba bean seeds are mixed with bio-fertilizer a commercial product that contains active microorganisms that are responsible for the atmosphere's fixation of nitrogen for 15 minutes. A 5.0 g soil sample was suspended in 95 ml of sterile distilled water, shaken, and heated to 75 oC for five minutes to create the soil solution [9]. Mention that the Nutrient Agar Medium (Sigma) was used to identify and purify Bacillus sp. The soil solutions were put on an agar plate medium after being serially diluted in sterile 0.85% NaCl solution from 10-1 to 10-6. For 48 hours, plates were incubated at 35 °C. On yeast extract glucose agar, several colonies were extracted during incubation. Bergey's Manual of Systematic Bacteriology was used to identify isolated bacteria using morphological, cultural, and biochemical methods [10].
Ammonium nitrate (33% N) was used as the nitrogen fertilizer; a 20 kg N/fed starting dosage was applied. Prior to the initial irrigation, 150 kg/fed of calcium super-phosphate (15.5% P2O5) and 25 kg/fed of potassium sulphate (48% K2O) were pre-sown. at 21 and 35 days following DAS seeding, in two equal dosages. Foliar applications of potassium silicate at the rates (0.0, 50, 75 and 100 ppm SiO3) occupy the sub plots.
Estimation at the heading stage
Leaf pigment content
The quantity of leaf greenness held by a plant was assessed using the Minolta-SPAD Chlorophyll Meter (Minolta Camera Co., Osaka, Japan). The SPAD-502 chlorophyll meter measures the absorbance of chlorophyll in the red and near-infrared spectrums to provide a numerical SPAD value that is proportional to the amount of chlorophyll in the leaf [11].
Relative water content
For every dry spell, measured the leaves' relative water content (RWC) [12]. For these ten mature leaves, five plants each plot (two leaves per plant) were selected at identical heights, and their fresh weight (FW) was recorded. The leaves were submerged in filtered water for 24 hours while being kept in low light, to calculate their weight. The leaves were dried at 75 °C for 48 hours to ascertain their dry weights (D.W.) after their turgescence weight (TW) was measured. The following formula was used to calculate RWC: RWC = (FW-DW)/ (TW-DW) × 100.
Determination of proline content
0.40 g of fresh plant material were homogenized with 1.25 cc of distilled water and then incubated for 30 minutes at 100 °C. After allowing the materials to cool to room temperature (22 °C), they were centrifuged for 10 minutes at 4000 rpm. Following the addition of 1.0 ml of a 1.0% solution of Ninhydrin in 60% acetic acid to the supernatant, the mixture was incubated at 100 °C for 20 minutes. After the samples cooled to 22 °C, 3.0 ml of toluene was added, and they were shaken and left in the dark for 24 hours to facilitate phase separation. state that a spectrophotometer was used to measure the absorbance of one milliliter of proline extract in a cuvette at a wavelength of λ = 520 nm [13].
Yield and its components
Samples were harvested from randomly selected plants in each experiment unit to estimate the average plant height (cm), pods length (cm) and 100 seed weight (g).
Biological yield:
The total biomass of the harvested plants (g/plant), then it was dried and weighted (g/plant).
Seed yield (g/plant), (ton/fed.)
It was obtained as the weight of clean seeds of the plot after threshing, and then it was transformed into tone per feddan.
Harvest index: Harvest index (H.I) calculated as follows:
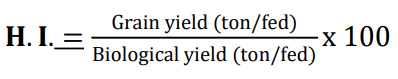
Phytochemical determination:
The nutrients content in Wheat grains (N, P, K, Na, Fe, Zu and Mn) were determined according to [14]. The crude protein percentage was extracted and determined by Macro-Kjeldahl method as described by [15]. The value of total crude protein was calculated by multiplying total values of total-N by factor 6.25. Total carbohydrate using the colorimetric method described [16].
Statistical analysis:
Homogeneity of variance between seasons was tested prior to combined analysis. Data were subjected to ANOVA appropriate for a split–split plot design using CoStat software (Version 6.4). Main effects and interactions were evaluated, and treatment means were compared using LSD at P ≤ 0.05 according to [17].
Results
Data presented in tables and figures represent the combined analysis of two successive growing seasons (2022/2023 and 2023/2024).
Effect of silicon
Physiological Traits
It’s clear from data in Table (3) revealed significant differences among silicon foliar application levels regarding chlorophyll content (SPAD), tissue water content (TWC%), and relative water content (RWC%).The application of 75 ppm Si recorded the highest values of chlorophyll (SPAD), TWC%, and RWC%, either with or without bio-fertilizer. The 100 ppm treatment ranked second, followed by 50 ppm, whereas the control treatment (0 ppm Si) recorded the lowest values for all studied physiological parameters.
Yield and yield components
Data presented in Table (4) showed significant differences among silicon treatments in plant height (cm), pod length (cm), and 100-seed weight (g). The 100 ppm Si treatment produced the tallest plants, longest pods, and heaviest 100-seed weight, regardless of bio-fertilizer application. Treatments were generally arranged in descending order as follows: 100 ppm > 75 ppm > 50 ppm > control. However, for plant height under bio-fertilizer application, 50 ppm ranked second, followed by 75 ppm. Results in Table (5) indicated significant variations in number of branches per plant, pod weight per plant (g), and number of pods per plant. The 100 ppm treatment significantly outperformed the other silicon levels, producing the highest number of branches, greatest pod weight, and highest pod number per plant, with or without biofertilizer. The descending order was 100 ppm > 75 ppm > 50 ppm > control. Regarding seed yield per plant and per fed(4200 m²), Table (6) demonstrated that the 100 ppm Si treatment produced the highest seed yield under both bio-fertilized and non-bio-fertilized conditions. The 75 ppm treatment ranked second, followed by 50 ppm, while the control recorded the lowest yields. The superiority of 100 ppm Si is consistent with its enhancement of all yield components, which was ultimately reflected in higher final seed yield per plant and per unit area
Macro and Micronutrients Components in Roots; Shoots and Seeds.
Data in Table (7) showed significant differences among silicon treatments in nitrogen (N %) content in roots, shoots, and seeds. The 100 ppm treatment recorded the highest N%, followed by 75 ppm, 50 ppm, and control. An exception was observed for seed N%, where control ranked third and 50 ppm fourth. Similarly, phosphorus (P%) content (Table 8) significantly increased with silicon application, with 100 ppm showing the highest values, followed by 75 ppm, 50 ppm, and control in descending order. Potassium (K %) content (Table 9) was significantly affected by silicon treatments. The highest values were generally obtained with 100 ppm and 75 ppm, followed by 50 ppm and control. Calcium (Ca %) content (Table 10) followed a similar pattern, where 100 ppm Si showed superiority over other treatments. Micronutrient contents in seeds (Fe, Zn, Mn, and Cu; Table (11) were also significantly influenced by silicon application. The 100 ppm treatment recorded the highest concentrations, followed by 75 ppm, 50 ppm, and control.
Effect of irrigation
Data presented in Tables (3–11) indicated that sufficient irrigation significantly outperformed deficit irrigation in all studied parameters. Under deficit irrigation (50% of water requirement), mean seed yield per plant with bio-fertilizer reached 145.2 g, representing 92.72% of that under sufficient irrigation (172.3 g/ plant). Without bio-fertilizer, deficit irrigation produced 109.7 g/ plant, equivalent to 89.77% of the sufficient irrigation treatment (122.2 g/plant). Similarly, seed yield per feddan under deficit irrigation reached 2.43 ton/fed with bio-fertilizer (89.01% of sufficient irrigation) and 1.97 ton/fed without bio-fertilizer (93.36% of sufficient irrigation). A comparable trend was observed for major yield components. Under deficit irrigation, pod weight per plant represented 95.15% (with bio-fertilizer) and 97.27% (without bio-fertilizer) of sufficient irrigation values. The number of pods per plant reached 90.55% and 96.12%, respectively, while 100-seed weight represented 96.67% and 97.41% compared to sufficient irrigation.
Interaction between silicon and irrigation.
The interaction between 100 ppm Si and sufficient irrigation showed significant superiority in most growth, yield, and nutrient parameters, including plant height, pod length, 100-seed weight, branches per plant, pod weight, pod number, seed weight per plant, seed yield (ton/fed), and macro- and micronutrient contents (N, P, K, Ca, Fe, Zn, Mn, and Cu). In contrast, the interaction of 75 ppm Si with sufficient irrigation recorded the highest values for physiological traits, namely chlorophyll (SPAD), TWC%, and RWC%.
Interaction among Silicon, Irrigation, and Bio-Fertilizer
The three-way interaction revealed that 75 ppm Si combined with sufficient irrigation and bio-fertilizer produced the highest chlorophyll content (SPAD), TWC%, and RWC%. However, the combination of 100 ppm Si, sufficient irrigation, and bio-fertilizer significantly enhanced plant height, pod length, 100-seed weight, branches per plant, pod weight, pod number, seed yield per plant and per fed, as well as N, P, K, Ca, Fe, Zn, Mn, and Cu contents, outperforming all other treatment combinations.
Proline Content:
Drought stress significantly increased proline accumulation at different growth stages, as illustrated in Figure (1). The highest proline content (mg g-¹ fresh weight) was recorded in untreated plants under water stress conditions. In contrast, foliar application of potassium silicate, particularly at 100 ppm Si combined with bio-fertilizer, resulted in the lowest proline content, indicating its effective role in mitigating water stress effects.
Fresh and Dry Weight:
Figures.(2and 3) illustrate the effects of potassium silicate and its interaction with bio-fertilizer on fresh and dry weight under sufficient and deficit irrigation. Sufficient irrigation significantly increased both fresh and dry weight compared to water stress conditions. The lowest values were recorded in untreated plants or those receiving 50 ppm Si without bio-fertilizer under stress conditions. Conversely, foliar application of 100 ppm Si combined with bio-fertilizer produced the highest fresh and dry weight under both irrigation regimes.
Discussion
The beneficial effects of silicon (Si) observed in the present study can be primarily attributed to its accumulation and deposition within plant tissues, particularly in epidermal and vascular structures. This deposition enhances cell wall rigidity and mechanical strength, thereby improving plant erectness and leaf orientation. Improved canopy architecture enhances light interception efficiency, which consequently increases photosynthetic performance and overall biomass accumulation. Moreover, strengthened tissues contribute to reduced pest susceptibility and improved structural stability, ultimately promoting plant growth and productivity. Several studies have demonstrated that the positive role of silicon under both biotic and abiotic stress conditions is associated with its capacity to mitigate multiple stress factors simultaneously. Silicon enhances plant water status by maintaining leaf water potential, improving stomatal conductance, sustaining photosynthetic activity, and preserving leaf erectness under high transpiration rates [18,19]. These physiological adjustments collectively contribute to improved stress tolerance and growth performance under deficit irrigation. In addition, silicon has been shown to maintain chlorophyll content and antioxidant defense systems under stress conditions. For instance, silicon application preserved chlorophyll concentration, glutathione (GSH), phenylalanine ammonia-lyase (PAL) activity, and vitamin C levels in tomato plants exposed to salt stress [20]. The enhancement of antioxidant metabolism reduces oxidative damage caused by reactive oxygen species (ROS), thereby protecting cellular membranes and photosynthetic machinery. The current findings indicate that foliar application of 100 ppm silicon combined with bio-fertilizer under deficit irrigation (50% of the recommended irrigation requirement) produced fresh, dry, and seed yield per plant, as well as seed yield per fed, reaching approximately 89–97% of the yield obtained under sufficient irrigation. These results suggest that silicon application can partially compensate for water limitation and significantly reduce irrigation water requirements without substantial yield penalties. From an agronomic perspective, this represents a practical strategy for improving water use efficiency under limited water resources. The observed yield improvement may be attributed to the growth-promoting effect of silicon, which enhances vegetative growth parameters and stimulates yield components. Previous studies have reported that increased yield following silicon foliar application is primarily associated with improvements in yield components. In cereals, these improvements included increased spike density, higher number of kernels per spike, and greater 1000-grain weight [21].
Similarly, reported that soil application of 1000 mg kg-¹ potassium silicate (K2SiO3) under deficit irrigation (50% field capacity) produced the highest biomass yield across four grass species [22]. Silicon-mediated drought resistance has also been linked to anatomical and hydraulic modifications. In rice, silicon enhanced drought resistance through silicification of the root endodis, which improved water uptake efficiency. Demonstrated that 50 ppm silicon increased stomatal conductance and transpiration rate, while alleviating water uptake reduction in hydroponically grown sorghum exposed to polyethylene glycol-induced water stress [23]. These improvements were attributed to silicon-induced enhancement of hydraulic conductance within plant tissues. Furthermore, osmotic adjustment plays a crucial role in plant adaptation to water stress. Increased proline accumulation, as reported in cotton plants under water stress, enabled maintenance of lower leaf water potential and facilitated continued water uptake from the surrounding [24]. The authors suggested that silicon application improved hydraulic conductance in leaves, thereby delaying the onset of osmotic adjustment at a given leaf water potential. Such mechanisms likely contributed to the improved physiological performance observed in the present study under deficit irrigation. Silicon is increasingly recognized as a beneficial element due to its regulatory role in enhancing plant tolerance to both salt and drought stress. Exogenous silicon application has been widely reported as an effective strategy for mitigating stress-induced growth inhibition [25]. Demonstrated that silicon improved drought tolerance in tomato plants by enhancing plant water status, osmotic regulation, photosynthetic efficiency, and nutrient balance, ultimately maintaining fruit yield [26]. Additionally, silicon deposition in epidermal cell walls reduces excessive water loss and limits uncontrolled transpiration, thereby improving drought tolerance [27]. Overall, the present findings are consistent with previous research and confirm that foliar application of silicon, particularly at 100 ppm combined with bio-fertilizer, enhances physiological performance, water relations, and yield stability under deficit irrigation conditions. These improvements appear to be mediated through structural reinforcement, enhanced photosynthetic capacity, improved antioxidant defense, and better hydraulic conductance. Therefore, silicon application represents a promising agronomic approach for sustaining crop productivity under water-limited environments.
Conclusion
Drought increasingly threatens global food security, demanding transformative strategies beyond conventional agronomy. This study reveals that the integration of silicon and biofertilizers acts as a synergistic, science-driven intervention that fundamentally enhances plant resilience under water-limited conditions. Silicon fortifies cell walls, reducing water loss, while biofertilizers activate root and metabolic adaptations together orchestrating a powerful, coordinated defense against drought stress. Far from incremental, this approach establishes a novel paradigm for climate-smart agriculture: scalable, resource-efficient, and capable of sustaining crop productivity under extreme environmental pressures. Future perspective: Unlocking the molecular and transcriptomic mechanisms underlying silicon–microbe synergy, combined with field-scale validation, will enable the translation of this integrative strategy into precision, climate-resilient agricultural systems a pivotal step toward securing global food production in the era of climate uncertainty [28-40].
Acknowledgement
The authors warmly thank the National Research Centre (NRC), Egypt. For completing this research honorably.
Competing interests
Authors have declared that no competing interests exist.
References
- Khalifa, R. (2019). Response of faba bean to alternate irrigation and cut-off irrigation combined with mineral phosphorus levels and biofertilizer at North Nile Delta soils. Egyptian Journal of Soil Science, 59(2), 175-191.
- FAOSTAT (2022). FAOSTAT.
- Ay, I., & Fayed, A. A. M. (2020). Response of dry seed yield of Faba bean “Vicia Faba, L.” to spraying with amino acids, organic acids,(NAA) growth regulator and micro nutrients. Alexandria Journal of Agricultural Sciences, 65(1), 7-16.
- Hungria, M., & Vargas, M. A. (2000). Environmental factors affecting N2 fixation in grain legumes in the tropics, with an emphasis on Brazil. Field crops research, 65(2-3), 151-164.
- Under Newly, S. F. B. V. (2011). Effect of row spacing on yield and its components of some faba bean varieties under newly reclaimed sandy soil condition. World Journal of Agricultural Sciences, 7(1), 68-72.
- Ali, A. M., Ibrahim, S. M., & Abou-Amer, I. (2019). Water deficit stress mitigation by foliar application of potassium silicate for sugar beet grown in a saline calcareous soil. Egyptian Journal of Soil Science, 59(1), 15-23.
- Gao X., Zou C., Wang L., &Zhang F. (2016). Silicon decreases transpiration rate and conductance from stomata of maize plants. J. Plant Nutr.29, 1637–1647. Gong
- Jackson, M.L. (1973). Soil Chemical Analysis. Prentice Hall, Ine., Englewood Cliffs, New Jersy, USA.
- Yörükçe, M. A., Aktas, B., Geroglu, Y., Poyrazoglu, E., & Biyik, H. H. (2017). Isolation and identification of bacteria from fruit garden soils in Aydin Province. International Journal of Secondary Metabolite, 4(2), 66-73.
- Bergey, D. H., Hendricks, D., Holt, J. G., & Sneath, P. H. (1984). Bergey's manual of systematic bacteriology. Vol. 2. Williams & Wilkins.
- Minolta, K. (1989). Chlorophyll meter SPAD-502 instruction manual. Minolta Co., Ltd., Radiometric Instruments Operations Osaka, Japan, 22.
- Castillo, F. J. (1996). Antioxidative protection in the inducible CAM plant Sedum album L. following the imposition of severe water stress and recovery. Oecologia, 107(4), 469-477.
- Filek, M., Biesaga-Koscielniak, J., & Dziurka, M. (2014). Determination of proline, carbohydrates and ethylene content and their role in drought stress in plant. Methodology of system approach to study drought tolerance in barley, Institute of Plant Genetics PAS, Poznan, 95-104, (2026).
- Cottenie, A., Verloo, M., Kiekens, L., Velghe, G., & Camerlynck, R. (1982). Chemical analysis of plants and soils. Lab. Agroch. State Univ. Gent, Belgium, 63, 44-45,(2026).
- A O A C (1990). AOAC Official Methods of Analysis. 15th Edition, Association of Official Analytical Chemists, Arlington.
- DuBois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., & Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Analytical chemistry, 28(3), 350-356.
- Williams, L. J., & Abdi, H. (2010). Fisher’s least significant difference (LSD) test. Encyclopedia of research design, 218(4), 840-853.
- Crusciol, C. A., Pulz, A. L., Lemos, L. B., Soratto, R. P., & Lima, G. P. (2009). Effects of silicon and drought stress on tuber yield and leaf biochemical characteristics in potato. Crop science, 49(3), 949-954.
- Shen, X., Zhou, Y., Duan, L., Li, Z., Eneji, A. E., & Li, J. (2010). Silicon effects on photosynthesis and antioxidant parameters of soybean seedlings under drought and ultraviolet-B radiation. Journal of plant physiology, 167(15), 1248-1252.
- Pinedo-Guerrero, Z. H., Cadenas-Pliego, G., Ortega-Ortiz, H., González-Morales, S., Benavides-Mendoza, A., Valdés-Reyna, J., & Juárez-Maldonado, A. (2020). Form of silica improves yield, fruit quality and antioxidant defense system of tomato plants under salt stress. Agriculture, 10(9), 367.
- White, B., Tubana, B. S., Babu, T., Mascagni Jr, H., Agostinho, F., Datnoff, L. E., & Harrison, S. (2017). Effect of silicate slag application on wheat grown under two nitrogen rates. Plants, 6(4), 47.
- Eneji, A. E., Inanaga, S., Muranaka, S. L. I. J., Li, J., Hattori, T., An, P., & Tsuji, W. (2008). Growth and nutrient use in four grasses under drought stress as mediated by silicon fertilizers. Journal of Plant Nutrition, 31(2), 355-365.
- Sonobe, K., Hattori, T., An, P., Tsuji, W., Eneji, E., Tanaka, K., & Inanaga, S. (2009). Diurnal variations in photosynthesis, stomatal conductance and leaf water relation in sorghum grown with or without silicon under water stress. Journal of plant nutrition, 32(3), 433-442.
- Gebaly, S. G., Ahmed, F. M., & Namich, A. A. (2013). Effect of spraying some organic, amino acids and potassium citrate on alleviation of drought stress in cotton plant. Journal of plant production, 4(9), 1369-1381.
- Hurtado, A. C., Chiconato, D. A., de Mello Prado, R., Junior,G. D. S. S., Gratão, P. L., Felisberto, G., ... & Dos Santos,D. M. M. (2020). Different methods of silicon application attenuate salt stress in sorghum and sunflower by modifying the antioxidative defense mechanism. Ecotoxicology and Environmental Safety, 203, 110964.
- Malik, M. A., Wani, A. H., Mir, S. H., Rehman, I. U., Tahir, I., Ahmad, P., & Rashid, I. (2021). Elucidating the role of silicon in drought stress tolerance in plants. Plant Physiology and Biochemistry, 165, 187-195.
- Mahmoud, L. M., Shalan, A. M., El-Boray, M. S., Vincent,C. I., El-Kady, M. E., Grosser, J. W., & Dutt, M. (2022). Application of silicon nanoparticles enhances oxidative stress tolerance in salt stressed ‘Valencia’sweet orange plants. Scientia Horticulturae, 295, 110856.
- Ahmed, F. F.; Mohamed, Y.& Abdalla, B. M. (2007). The relation between using some antioxidants and productivity of Sewa date palms. Minia J. Agric. Res. Dev. 27 (4), 753-770, (2026).
- Mohamed, A. A., Eissa, M. A., & Bakheit, B. R. (2024). Algal and nanoparticles of silicon and boron foliar application efficiency for maximizing yield, and quality traits of two faba bean varieties. Egyptian Journal of Agronomy, 46(1), 213-227.
- Crépon, K., Marget, P., Peyronnet, C., Carrouée, B., Arese, P., & Duc, G. (2010). Nutritional value of faba bean (Vicia faba L.) seeds for feed and food. Field crops research, 115(3),329-339.
- Das, K. K., Swamy, G. S., Biswas, D., & Chnaniya, K. K. (2017). Response of soil application of diatomaceous earth as a source of silicon on leaf nutrient status of guava. Int. J. Curr. Microbiol. App. Sci, 6(4), 1394-1399.
- Dawood, M. G., Abdel-Baky, Y. R., El-Awadi, M. E. S., & Bakhoum, G. S. (2019). Enhancement quality and quantity of faba bean plants grown under sandy soil conditions by nicotinamide and/or humic acid application. Bulletin of the National Research Centre, 43(1), 28.
- El-Kramany, M. F., Bahr, A. A., Manal, F. M., & Kabesh, M. O. (2007). Utilization of bio-fertilizers in field crops production 16-groundnut yield, its components and seeds content as affected by partial replacement of chemical fertilizers by bio-organic fertilizers.
- Epstein, E. (1999). Silicon Annu. Rev. Plant Biol. 50,641-664.
- Epstein, E. &A.J. Bloom. (2005). Mineral nutrition of plants: principles and perspectives. 2nd ed. Sunderland (MA): Sinauer Associates, Sunderland, MA. (2026).
- Gomaa, F. S. (2025). SUSTAINABLE FABABEAN (Viciafaba L.) USING VERMICOMPOST AND NITROGEN-FIXINGBACTERIA (Rhizobium sp.). Zagazig Journal of Agricultural Research, 52(1), 1-19.
- Fouda, K. F. (2017). Effect of phosphorus level and some growth regulators on productivity of faba bean (Vicia faba L.). Egypt. J. Soil Sci, 57(1), 73-87.
- H., Chen K. Chen G., Wang S. &Zhang C. (2016). Effects of silicon on growth and yield of faba bean under drought. J. Plant Nutr.26 (5):1055-1063.
- Shaaban, M. M., & Abou El-Nour, E. Z. A. (2014). Macro and Micro-Nutrients Concentrations and Uptake by Maize Seedlings Irrigated with Fresh or Saline Water as Affected by. Am J Plant Physiol, 9(3), 95-102.
- Sadras, V. O., Villalobos, F. J., Orgaz, F., & Fereres, E. (2017). Effects of water stress on crop production. In Principles of agronomy for sustainable agriculture (pp. 189-204). Cham: Springer International Publishing.