
Insights of Herbal Medicine(IHM)
ISSN: 2834-7749 | DOI: 10.33140/IHM



Research Article - (2024) Volume 3, Issue 2
Effect of Salinity and Humic Acid on Growth, Biomass, and Yield of Mung Bean (Vigna Radiata L.) Cultivars
2School of Environment and Sustainable Development, Central University of Gujarat, Gandhinagar, India
Received Date: May 20, 2024 / Accepted Date: Jun 10, 2024 / Published Date: Jun 24, 2024
Copyright: ©©2024 Indra Jeet Chaudhary, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation: Patankar, A. V., Nigam, B., Chaudhary, I. J. (2024). Effect of Salinity and Humic Acid on Growth, Biomass, and Yield of Mung Bean (Vigna Radiata L.) Cultivars. Insights Herbal Med, 3(2), 01-14.
Abstract
Higher salinity affects 20% of farmed land and 33% of irrigated land area globally. These affect causes food crises worldwide. Humic acid provide protection against stress and promote the plant growth and productivity. Therefore, this study focuses on dose response relationship between exogenous applied humic acid and salt stress on Mung (Vigna radiata L.) cultivars. Resultants theapplication of humic acid under salt stress improvedthe plant growthand yield of selected cultivars. The higher salinity (100 ppm) caused higher effect on both selected cultivars than 50ppm salt as compared to control plants. While applied protectants 40%HA with 50 ppm salt is more effective than 40%HA with 100ppm salt in both selected cultivars. Based on biomass and yield reduction Pusa Baisakhi is least sensitive to salt than ML-131. Study also concise that salt stress depends on growth stages of plant and higher biomass reduced at 20 DAS in cultivar Pusa Baisakhi (-11.15%) and at 40 DAS in cultivar ML-131 (-25.62%). The higher production of yield due to application of humic acid was seen (65.95%) in cultivar ML-131 than cultivar Pusa Baisakhi (8.54%). Therefore, the study promoted that under salt stress conditions humic acid uses as a protectant and cultivar Pusa Baisakhi as a tolerance species will be useful tool for sustainable agriculture.
Keywords
Mung Bean, Salt Stress, Humic Acid,Tolerance Plants, Sustainable Agriculture
Introduction
World wide population reachesto 9.6 billion will effectively necessitate an improvement in agricultural productivity for an additional 2 billion people over the time [1]. The sustainable supply of food for the rising population around the world, may be impacted by the salinity of arable land [2]. In significant cereal crops similar to maize, rice, and wheat, salinity has been associated to rising yield losses. Salinity in the soil is the major factor for degradation of the land. According to statistics, the total land area affected is approximately 1125 Mha, of which 76 Mha are damaged by salinization and sodification caused by humans [3-6]. The continuation of soil salinization at current rate, the most arable land will disappear by 2050. According to study by, the amount of irrigated land used for agriculture worldwide is likely to rise by more than 40% in the near future, accounting for about 45 million hectares (20% of the irrigated land) and producing 33% of the world's food. In India, salt affects over 75% of the soil [7,8]. According to, Gujarat (2.23 Mha) has the most affected land area, followed by Uttar Pradesh (1.37 Mha)> Maharashtra (0.61 Mha)> West Bengal (0.44 Mha)> and Rajasthan (0.38 Mha) [9].
The biggest difficulty in the future years will be controlling crop productivity on salty soil. Plants under salinity stress undergo morpho-physiological and biochemical changes that result in membrane damage, ineffective photosynthesis, unbalanced nutrient levels, changes in the level of growth regulators, enzyme inactivation, and metabolic inhibition [10]. According to, the detrimental effects of salt stress also lead to a larger formation of ROS by upsetting the redox balance in plant cells, which damages vital cellular molecules and organelles. The production of osmoprotective compounds, stomatal regulation for maintaining water status in salinity-stressed plants, and enzymatic and non-enzymatic antioxidants are just a few of the defence mechanisms that plants naturally develop [2,11,12]. However, in extreme circumstances of salinity stress, defence mechanisms are weakened, and the plants experience the negative effects of damage brought on by salinity.
Ongoing research is being done to determine the best methods for developing salt resistance in plants, which will eventually enable them to counteract the damaging effects of salinity. According to recent research, a variety of growth controllers, including hormones that have been used to prime plants, appear to have promising methods to prevent salt toxicity and increase crop output [13,14]. Being one of the world's top producers of oilseeds and leafy vegetables, India plays a crucial role in the country's agricultural economy. One of the most significant edible legume crops is the mung bean, which is cultivated on more than 6 million hectares globally (or roughly 8.5% of the world's pulse area) and is consumed by the majority of Asian families. Mung bean contains 48% Protein and 21% Carbohydrates. While mung bean is short duration horticulture of the region. They're also high in iron, protein and fibres, rich in vitamins and minerals. But when fertile soils get more salinized, these key crops become less productive, which has an impact on farmers' capacity to remain economically viable.
A class of chemicals known as humic acid binds to and facilitates the uptake of nutrients and water by plant roots. states that yields can be significantly increased by high humic acid levels [15]. The humic material has a larger molecular weight than fulvic acid and is only soluble in water at higher soil pH values. In intact soil, humic acid can persist for decades. Adding organic material is definitely a great technique for improving a leached soil or one that doesn't respond chemically, like specific sands. Nutrient cations are hard for sand with a poor cation exchange capacity (CEC) to hold onto; as a result, these charged particles can quickly seep deeply into the soil and become inaccessible to plants. They transport the water and nutrients the plant needs when they reach the roots.Humic acid is a compound that has been found to promote plant growth by raising the chlorophyll levels, the processes of photosynthesis rates, and the formation of roots. Larger plants, quicker development, and bigger production may come from this strategy.Increased soil fertility and encouraging the development of good bacteria and fungi, which degrade organic material and release nutrients to plants.Humic acid has been found to improve the capacity of plants to withstand environmental stresses including heat, cold, and drought. In general, high humic acid levels can boost the fertility of the soil, stimulate plant growth, improve the structure of the soil, and increase accessibility to nutrients stress tolerance.
Therefore, the presented study was conducted to measure the protective role of humic acid on mung bean cultivars under salt stress. Despite that, study also conducted for assessment of intraspecific sensitivity of Mung bean cultivars.
Methods
Study Site
Apot investigation was carried out atDepartment of Environmental Science, Savitribai Phule Pune University, Maharashtra, India.Pune District occupies an area about 411 acres, contains black soil where the main crop is paddy. In Pune, the dry season is primarily clear, the wet season is oppressive, windy, and cloudy, and the weather is hot all year round. Pune is located in the western part of India and has a semi-arid environment with warm summers, monsoon season, and cool winters. During February to April, which is the transition period from winter to summer, the temperature in Pune typically variates from 20°C (68°F) to 35°C (95°F).
Plant Materials
Healthy and genetically homogeneous seeds of two Mung bean (Vigna radiata L) cultivars, Pusa Baisakhi and ML-131, were chosen for the pot experiment. Crop cultivars were chosen based on their availability and suitability at the experimental region. Mung bean is commercially important and widely grown crop that are cultivated all around the world. Though, both are diverse in their lifecycle. Traditionally, mung bean has been grown during kharif season. The seeds of the experimental crops were procured from Agriculture College, Pune.
Experimental Design and Treatments
The pot experiment was conducted for assessment of salt effect and role of humic acid on growth, development yield of mung bean cultivars. Twenty genetically uniform seeds of Mung bean (Pusa Baisakhi and ML-131) were sown in 10 × 20 cm sized clay pots.Each pots filled with 10 kg of soil and also mixed with 250 g vermicomposting and irrigated regularly from seed germination to till harvesting. The used soil was sandy loam with 7.4 pH and 0.50 dSm− 1 electrical conductivity (EC). Mung bean, is a warm-season crop that requires temperatures ranging from 25 to 35 degrees Celsius for maximum growth and development. Temperatures exceeding 35°C or below 15°C might have an adverse effect on its development and output. Mung is best cultivated in India during the summer season when the weather is warm and favourable for growth. It is important to note that the exact temperature range during Mungbean cultivation might vary based on a variety of factors such as location, season, and weather conditions. HA (Humic Acid) at concentration of 40% were poured every 10thdays of intervals at morning from germination to maturity. A control group (without saltand humic acid treatment) was also kept for comparison. Treatments used in the experiment included T1 (control), T2 (50 ppm NaCl), T3 (100 ppm NaCl), T4 (50 ppm NaCl + 40%HA), and T5 (100 ppm NaCl + 40%HA). The data presented are the average of three concordant readings from the experiment, which was run in triplicate.
Sampling and Plants Analysis
Plants were taken for growth and biochemical analysis at juvenile and vegetative stages (20 and 40 days after sowing). Yield parameters were analyses after harvesting of plants.
Growth and Biomass Analysis
The root and shoot lengths were measured using a metre scale and reported as cm plant-1. The leaf area was measured graphically. For this purpose, plant leaves were laid out on graph paper, and the area covered by each leaf was measured and reported as cm2 plant-1. The plant components were thoroughly cleaned, dried with tissue paper, and then separated. They were then weighed again and maintained in a hot air oven at 80°C until a steady dry weight was reached.
Growth Indices
Hunt (1982) described a formula for measuring biomass accumulation and allocation.
Root: Shoot ratio (g g-1) = RW / SW
Where, RW = root dry weight; SW = shoot dry weight.
Determination of RWC
Firstly,fresh weight (fw) of leaf was measured and after that same leaves immersed in distilled water for 24 h. then after 24 h turgid weight (tw) was achieved and, the same tissue was oven dried for 48 hours at 80°C for dry weight (dw). The RWC was calculatedby [16].
formulae:
fw − dw
RWC =--------------------------- × 100
tw − dw
Estimation of Membrane Permeability
Membrane permeability was measured as electrolyte leaked out from fresh leaves. For this, twenty discs (1 cm2) of fresh leaves were cut by punching machine. These discs were put into test tubes with 10 mL of distilled water in each. A conductivity meter was used to measure the electrolyte leakage in the solution after two hours at room temperature.
Biochemical Analysis
Total Chlorophyll and Carotenoids
Leaf sample (0.1 g) wascrushed with 10 mL of 80% acetone. The crushed solution kept overnight at 4 0C. Then after centrifuged OD (optical density) of the solution was opted at 663 nm and 645 nm. Total chlorophyll and carotenoids were calculated following the formula described by [17,18].
Determination of Flavonoid Content
The Flavonoid Content was established using Flint method [19]. 100 ml extraction solution containing ethanol and acetic acid (99:1, v/v) was used with 0.1 g of fresh leaf. After two minutes boiling the solution cooled and centrifuged at 8000x for 15 minutes, and OD taken at 250–350 nm. The number of flavonoids presented(A mg− 1 fresh weight).
Estimation of Ascorbic Acid
Based on the reduction of 2, 6-dichlorophenolindophenol (DCPIP), the ascorbic acid concentration was extracted and estimated using thetechnique[20]. Using the following formula, a standard curve made from an aqueous solution of ascorbic acid was used to compute the total amount of ascorbic acid:
Ascorbic acid (mg g− 1fresh leaf) = [{Eo− (Es− − Et)} × V] / (v × W × 1000)
Analysis of Yields Characteristics
Yield characteristic was measured with the help of number of seed (plant-1), number of pods (plant-1), seed and pods weight (g/ plant-1). The following formulas were used to determine the harvest index using total plant biomass. (Economic yield / Biological yield) x 100 is the harvest index (HI %). 1000 seed weight was used to calculate the test weight.
Statistical Analysis
Five treatments were included in the randomized block design of the trial. Using Origin Pro 2023 software, principal component analysis (PCA) was applied to the data obtained for each parameter. Using Duncan's Multiple Range Test by SPSS (SPSS Inc., version 17.0), the least significant difference was computed at 0.05% probability level to identify the significant differences among the mean values.
Results
Soil salinity negatively affected growth, development of mung bean cultivars used for the presented study however; exogenous protectants humic acid were found to induce salt tolerance in tested cultivars under salt stress conditions.
Plant Morphological Characters
The results indicated that the reduction of leaf area in experimental cultivars were increased with increased salt concentration (Figure. 1). Among the cultivars, ML 131 exhibited the greatest decrease in leaf area (-22.61%) at 20 days after sowing (DAS) under treatment T3 (100 ppm salt), compared to the control (T1). Conversely, cultivar Pusa Baisakhi displayed the least reduction in leaf area (-15.09%) at 20 DAS in the T3 treatment, as compared to the control plants (T1). On the other hand, the application of humic acid externally resulted in an increase in leaf area for the selected cultivars, with the highest enhancement observed in cultivar ML 131 (53.57%) at 20 DAS in treatment T5.Based on the results, Pusa Baisakhi exhibited a control root length of 4.3 cm in the juvenile stage. However, a decrease in root length was observed when both humic acid and 100 ppm salt stress (T3) were applied. Conversely, in the vegetative stage, treatments T4 (9.7 cm) and T5 (9.5 cm) demonstrated a significant increase in root length, indicating that the use of exogenous protectants like humic acid could potentially promote root development and growth, particularly under salt stress conditions. Throughout the experiment, the ML 131 cultivar consistently displayed robust development. These findings suggest that the application of exogenous protectants may have stage-specific effects on root growth, which could be influenced by factors such as cultivar selection and timing. Additionally, it was observed that the Pusa Baisakhi cultivar exhibited a significant increase in branch length compared to the ML 131 cultivar, both in the juvenile and vegetative stages. Notably, treatment T4 demonstrated the highest growth, with the presence of exogenous protectants contributing to this outcome.Based on the results, no significant difference in total plant height was observed among the treatments during the juvenile stage. However, in the vegetative stage, all treatments exhibited a notable increase in plant height compared to the juvenile stage. Conversely, the ML-131 variety consistently displayed a significant decrease in plant height throughout both the juvenile and vegetative periods (Fig.1).
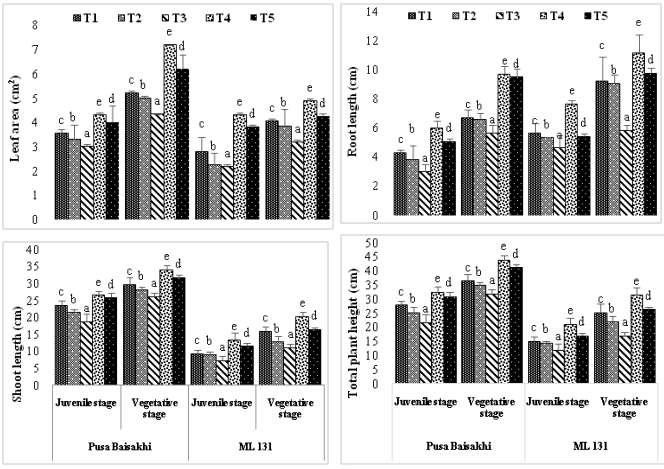
Figure 1: Effect of salt and humic acid on leaf area (cm2 plant-1), Shoot height (cm plant-1), root height (cm plant-1) and total height (cm plant-1) of selected cultivars of Mung bean (Mean ± standard deviation of three replicates presented by thin vertical bars, Value within the each column same letter represent not significantly different (p< 0.05) using Duncan’s Multiple Range Test).
Plant Biomass Characters
The leaf fresh weight was significantly impacted by the salt stress; treatment T3 (100 ppm salt) showed a greater drop in leaf fresh weight than the control plant (T). Treatment T4 (23.67%) had the highest fresh leaf weight in Pusa Baisakhi. The results indicate that the application of external protective chemicals positively affects the turgid weight of leaves and encourages Pusa Baisakhi plants to develop more vegetative. Treatment T4 showed the most notable improvement in leaf turgid weight for both Pusa Baisakhi and ML-131 cultivars (23.90% and 60.62% respectively).The results indicate that the addition of 40% humic acid had a positive impact on the growth of Pusa Baisakhi during its juvenile stage, with a significant increase observed in treatment T4 (Fig.2). However, this impact was not observed in the ML-131 variety, which did not show significant growth in response to the addition of humic acid.
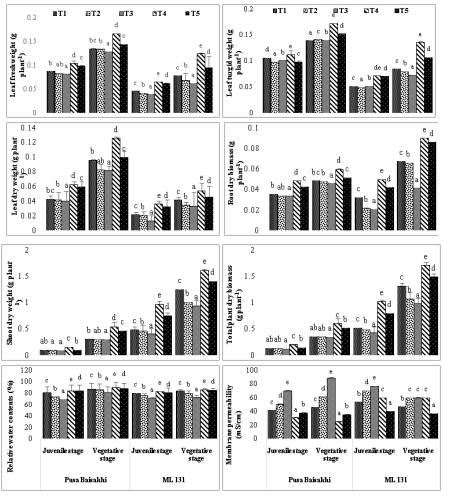
Figure 2: Effect of salt stress and humic acid on leaf fresh weight (g plant-1), leaf turgid weight (g plant-1), leaf dry weight (g plant-1), root dry weight (g plant-1), shoot dry weight (g plant-1), and total plant dry weight (g plant-1), relative water content (%) and membrane permeability (mS/cm) of selected cultivars of Mung bean (Mean ± standard deviation of three replicates presented by thin vertical bars, Value within the each column same letter represent not significantly different (p< 0.05) using Duncan’s Multiple Range Test).
In the juvenile stage of Pusa Baisakhi, the untreated Moong bean plants in treatment T3 exhibited a decrease in underground growth metrics such as root biomass, shoot biomass, and total plant dry biomass. However, ML131 cultivar showed enhancement in all these parameters compared to the other cultivars. The application of treatments at 20- and 40-day intervals resulted in changes in growth characteristics.The findings demonstrate a notable distinction in root and shoot length between the juvenile stage of Pusa Baisakhi and ML-131 in the control stage (T1). This variation is likely attributable to genetic differences between the two cultivars. Moreover, during the vegetative stage, both Pusa Baisakhi and ML131 exhibit a similar pattern in root and shoot length, with the lengths of the root and shoot being comparable. These results indicate that both genetic and environmental factors play a role in the development and growth of root and shoot systems in these cultivars. Further research is necessary to explore the underlying mechanisms behind these observations.
RWC and Membrane Permeability
One helpful metric for determining a plant's physiological water status is Relative Water Content (RWC). Under both salinity levels, the results showed a significant drop in RWC for all experimental crops (Fig. 2). Salinity had a greater effect on RWC in the ML131 cultivars than in Pusa Baisakhi. Humic acid addition attenuated for the salinity-induced improvement in RWC. However, the treatment T4 and T5 helped in mitigate the reduction in RWC caused by salinity stress. Notably, the compensatory effect of T4 treatment was more pronounced than that of treatment T5.Increased salinity levels resulted in higher electrolyte leakage, indicating increased membrane permeability due to salt stress. The severity of salinity-related membrane damage was more pronounced at higher salinity treatment levels, particularly at 100 ppm. Among the moong cultivars, Pusa Baisakhi exhibited the greatest increase in electrolyte leakage. Exogenous application of humic acid proved to be more effective in reducing electrolyte loss at lower salinity levels. However, interestingly, the utilization of 40% humic acid had a negative impact on the development of Pusa Baisakhi in both the juvenile and vegetative stages, contrasting with its effects on other cultivars.
Biochemical Changes
Mung bean cultivars' chlorophyll content was negatively impacted by soil salinity, particularly at higher salt concentrations (Fig. 3). Exogenous protectants, on the other hand, increased the amounts of photosynthetic pigments under both salinity levels, but at a slower rate in the presence of higher soil salinity. Treatment T4 plants showed a greater trend in total chlorophyll content, chlorophyll a, and chlorophyll b than T5 and control plants. When subjected to T4 treatment, the Pusa Baisakhi cultivar showed the greatest increase in total chlorophyll content when compared to the ML-131 cultivar. It's crucial to remember that although total chlorophyll levels increased, the concentration of chlorophyll a did not significantly rise during this growth period. This suggests that the increase in chlorophyll levels was primarily driven by higher levels of chlorophyll b or other forms of chlorophyll.
The results suggest that the application of humic acid significantly affects the carotenoid levels in both Pusa Baisakhi and ML131 cultivars. Specifically, the plants exposed to salt stress alone did not exhibit comparable growth, but the addition of humic acid increased carotenoid levels and promoted growth. These findings suggest that humic acid may enhance nutrient absorption and plant development under adverse conditions.During the juvenile stage, both Pusa Baisakhi and ML131 cultivars exhibited similar growth rates based on carotenoid level analysis. However, treatments T1, T4 (40% humic acid + 50 ppm NaCl), and T5 (40% humic acid + 100 ppm NaCl) showed a significant increase in carotenoid levels at 40 days after sowing (DAS) for both cultivars. The rise in carotenoid levels in the treated groups suggests that humic acid may play a beneficial role in carotenoid production.Ascorbic acid was decreased by salinity stress in all the ages in both cultivars (Figure:3).However, exogenously applied humic acid enhanced this antioxidant under both level of salinity. Treatment T4 was found more effective on this antioxidant than T5. Maximum increase of ascorbic acid was observed in cultivar ML131 (14.28%)at 40 DAS under T4 treatment.
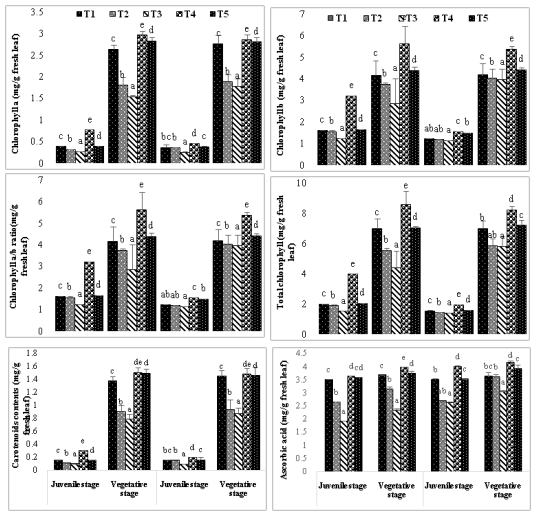
Figure 3: Effect of salt and humic acid on chlorophyll a (mg/g fresh leaf), chlorophyll b (mg/g fresh leaf), chlorophyll a/b ratio (mg/g fresh leaf), total chlorophyll (mg/g fresh leaf), carotenoids contents (mg/g fresh leaf) and ascorbic acid contents (mg/g fresh leaf) of selected cultivars of Mung bean (Mean ± standard deviation of three replicates presented by thin vertical bars, Value within the each column same letter represent not significantly different (p< 0.05) using Duncan’s Multiple Range Test).
Yields Characteristic
The opted results indicated that under both levels of salinity, there was a significant decrease in yield, number of capsules, and number of seeds, seed weight, and total yield of the plants across all ages for both cultivars (Figure:4). This suggests that salinity negatively impacted the overall productivity of the plants. Additionally, similar trends were observed in the harvest index and test weight of the plants, with a decrease in these parameters under salinity stress. This implies that the quality and efficiency of the plants' yield were also compromised under salinity conditions.
However, the application of humic acid as an exogenous treatment showed promising results in enhancing yield even under salinity stress. Specifically, Treatment T4, which likely involved a particular concentration or application method of humic acid, was found to be more effective in promoting yield compared to Treatment T5. This suggests that the specific application protocol or concentration of humic acid can influence its effectiveness in mitigating the negative effects of salinity on plant productivity. Maximum increase of total yield was observed in cultivar ML-131 (65.95%) at under T4 treatment.
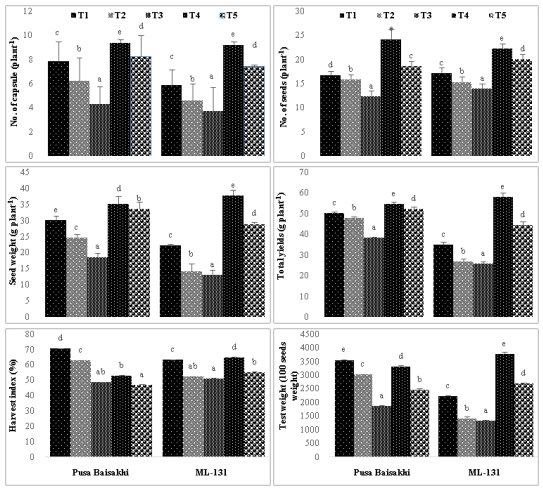
Figure 4: Effect of salt and humic acid on No. of capsule (plant-1), No. of seeds (plant-1), seed weight (g plant-1), total yields (g plant-1), harvest index (%) and test weight (100 seed weight) of selected cultivars of Mung bean (Mean ± standard deviation of three replicates presented by thin vertical bars, Value within the each column same letter represent not significantly different (p< 0.05) using Duncan’s Multiple Range Test).
Statistical Analysis
Principal component analysis (PCA) is used to summarize the data set and description of the multiple variables. It’s useful for highly correlated data particularly variables within the data set. The present study summarized the relationship between treatment, age, and cultivars, and variation was given below.PCA analysis of mung bean cultivar Pusa Baisakhi shows the percentage of variance bi-plot PC1 83.36% and PC2 13.05% with Eigenvalue PC1 13.33 and PC2 2.08. Correlation between parameters of cultivar Pusa Baisakhi showed a strong relation in leaf area, root, shoot length, plant height, leaf fresh dry weight, total biomass, carotenoids, total chlorophyll, seed weight and test weight at PC 1 with loading plot value 0.26 while total yield and harvest index are slightly correlated with loading value 0.25. The membrane permeability of Pusa Baisakhi shows negative loading value (-0.09) at PC 1. Treatment wise higher negative score value (-1.33) was seen in treatment 100ppm salt stress than 50ppm salt score value (-1.06). HAtreated plants show positive correlation at PC1 and higher score value was found in 40%HA+50ppm salt (1.61). Age wisehigher values was seen at vegetative stages than juvenile stages (Figure:5A). PCAs analysis indicate that the Pusa Baisakhi cope up their growth at vegetative stage.Cultivar ML-131 was also following the similar trends with age and treatments. The percentage varianceon both PCs are noted higher at PC1 81.85% with eigenvalue 13.09 and PC2 (11.06%) with eigenvalue 1.76. Shoot length, plant height and biomass of cultivar ML-131 showed loading value 0.27 and was highly correlated with each other’s. While root length, leaf fresh weight, leaf dry weight, seed and test weight was slightly correlated with above parameters with loading values (0.26).Carotenoid contents, total chlorophyll and total yield showed and similar group of correlation with loading value 0.25 while leaf area and harvest index showed 0.24 loading value.Score plot of PC1 with treatments showed positive values at vegetative stages of plants and negative values shown in all treatment at juvenile stages (Figure:5B).


Figure 5: Principle component analysis (PCA) correlation bi-plot of growth, biomass and yield response to salt stress and humic acid treatment. Symbol represent the standardized scores on PC1 (x-axis) and PC2 (y-axis) for the salt stress and humic acid treatment of Mung bean (cv. Pusa Baisakhi and ML-131). Vector coordinates represent the correlations between standardized variables and principle components (PCs).
Discussion
The influence of salinity on plant development, metabolism, and yield loss is significant and multifaceted. In the present research, the influence of salinity stress on mung bean cultivars has been evaluated. Numerous external protectants have been suggested to lessen the impacts of salinity on plants. Consequently, this study also investigates the dose response relationship of mung bean cultivars when treated with exogenous protectants. The relations between protectants and salt effect on cultivars is discussed below. Several researchers have reported reduced growth in numerous plant species under soil salinity circumstances [5,20,21]. The current study also observed decreased growth in mung bean cultivars, including shoot, root length, leaf area, and biomass, due to salinity. The degree of growth reduction was found to be higher with increasing salinity treatment levels. Acosta-Motos highlighted that growth retardation plays a crucial role in determining the extent of salinity-induced damage, irrespective of the plant species [20]. Reduction in leaf area is a significant adaptation mechanism for coping with salt stress in numerous crops, including halophytes, leading to a decline in the assimilatory unit of the plant and reduced water consumption [22,23]. Shahidproposed that physiological responses, such as water status regulation, stomatal behaviour, ion balance, mineral nutrition, and photosynthetic efficiency, are responsible for the decline in plant growth [20]. Elâ?Shafeyreported that the idea, stating that the inhibition of cell division and cell elongation under salt stress is the primary cause of reduced growth, biomass, and yield [24]. These findings are consistent with the results of the current study, which observed lower plant height and leaf area in mung bean cultivars grown under salinity. The exogenous protectant, Humic acid, used in this study, is renowned for promoting growth in various crops. The present study also observed improved growth with application of protectants under salt stress. In the current study, salinity considerably decreased the plant height and dry biomass of both mung bean cultivars. However, foliar humic acid spray prevented the detrimental effects of salt stress by promoting the development of the mung bean plants in a salt-stressed environment. The fact that humic acid comprises a number of vital elements needed for typical plant growth may be the cause of the humic acid-induced growth enhancement [25]. Numerous additional studies have also demonstrated the positive benefits of humic acid on the growth of other crops produced in challenging conditions. For instance, according to a study by the application of humic acid at a concentration of 25 kg ha-1 improved the quality and development of maize fodder in a number of different circumstances [26]. Increases in protein synthesis, enzymatic activity, respiration, and photosynthetic pigments may be the reason of the growth stimulation that HA induced in plants [27].
Under increasing salinity stress, the fresh and dry weight of leaves, shoots, and roots decrease, which is consistent with previous studies showing reduced plant fresh and dry mass under salinity stress [28-29]. Lessening in dry weights under salinity stress may be attributed to the inhibition of reserved food hydrolysis and its transfer to the growing stem [30]. Salt also affects the size of final cells and the rate of cell synthesis, thus impacting the overall mass of the plants [31]. In the present study, salt treatments significantly decreased the dry weight of leaves, shoots, and roots in mung bean cultivars, consistent with the findings of Ali [32]. However, the application of the exogenous protectant humic acid increased the dry weight of leaves, shoots, and roots[33]. In presented study applied humic acid increased the growth and biomass of selected plants species.Relative water content (RWC) is measured of plant water content, reflecting the metabolic activity in plant tissues. Salt stress regularly disrupts the water balance in plants, leading to a significant decrease in RWC as salinity levels increase. This reduction in RWC under salinity stress can be attributed to the diminished root system, which becomes less efficient in compensating for water harm through transpiration owing to a reduction in the absorbing surface [34,35]. The higher salinity levels, did not show significant differences in water content of the plant, but there was a sharp decrease in plant height, leaf area, and root length. Consistent with a study, RWC was found to be reduced with increasing salinity levels[36]. In the presented study, the application of humic acid resulted in increased root growth in the mung bean cultivars, leading to higher RWC and improved metabolic activity.
One of the main difficulties to sustainable agriculture is salinity, which reduces plant productivity worldwide by compromising a variety of physiological, biochemical, and molecular processes [37]. The membrane permeability is the factor that affect plant growth.Electrolytes are released from the cell when the plasma membrane loses its integrity. This phenomenon is typically observed under various stress conditions, including salinity, ozone, metals, UV-B etc. [28,38-41]. The reduction of water content in cellular system caused membrane permeability. In presented study also seen that higher reduction of relative water content and higher membrane permeability in salt stress condition plants.Stresses in the environment have a major impact on the amounts of photosynthetic pigments, which are essential for plants to absorb energy [42]. A vital component of photosynthesis, chlorophyll is essential to a plant's ability to endure [43]. Plants use photosynthesis to transform solar energy into chemical energy for food production [44]. Salinity, mainly through stomatal closure, leads to a reduction in the photosynthetic rate [45]. Plants exposed to salinity experience a drop-in chlorophyll content, which is thought to be a sign of oxidative stress caused by the inhibition of chlorophyll production and the activation of chlorophyllase enzyme destruction [46]. The reduced chlorophyll levels suggest the existence of a photoprotection mechanism that either slows down synthesis or accelerates breakdown to lower light absorbance [47]. In the present study, salt treatment resulted in a considerable reduction in photosynthetic pigments in mung bean cultivars. High salinity levels had a more significant impact on chlorophyll loss compared to low salinity levels. A related study by also observed a decrease in chlorophyll content in spinach and soybean cultivars as salinity increased [31]. While in presented study application of humic acid to salinity stressed mung bean cultivars boosted photosynthetic pigments. Similar findings were obtained by El-Sarkassyin pepper plants and Ennabin Mexican Lime Trees [48-49]. The humic acid raised photosynthetic pigment may be enhancing the absorption of nutrients in plants [50].
The organelles responsible for the manufacture and storage of carotenoids in plant cells are called plastids, and they contain antioxidant and photoprotective qualities [51]. Carotenoids serve two essential roles in plants. They act as pigments that absorb light energy for photosynthesis and are crucial for protecting photosystems from photo-oxidation. The reduction in carotenoid content indicates that the protective function of carotenoids may not be one of the primary mechanisms under salinity stress. However, in the current study, the treatment of mung bean cultivars with humic acid enhanced the carotenoid content even under salinity stress. El-Ghamry showed that application of HA increase carotenoid content in faba beans [52]. While salt application reduced carotenoids contents in both selected cultivars.One well-known non-enzymatic antioxidant that is essential for protecting plants from oxidative damage is ascorbic acid (AsA). It improves the redox state of cells. According to research by Akram plants with completely formed chloroplasts and mature leaves had higher ascorbic acid contents [22]. In another study, Caverzan explained that under stressful conditions, protein ascorbate peroxidase utilizes ascorbic acid to balance H2O2 levels in plants [53]. However, the mung bean cultivars in the presented study showed lower ascorbic acid levels under salt stress. An earlier work by similarly noted this drop in AsA content during saline stress (2014) [54]. The decrease in AsA production and regeneration in plants under salinity stress was caused by the decreased activity of the AsA-regenerating enzymes, MDHAR and DHAR. Conversely, higher AsA levels in the presence of protectant might be attributed to improved activity of AsA-regenerating enzymes or reduced degradation caused by salt stress.
Agricultural productivity serves as the backbone of the global economy therefore production of crop yield is an important task for formers. Now a day’s environmental stresses increased due due industrial development and caused negative impact on plants and agricultural crops too. The crop productivity is significantly affected by various environmental stresses, leading to yield losses. One of the major concerns in agriculture is the reduction in yield caused by environmental stresses, including salinity [31,55]. The decrease in yield can be attributed to reduced photosynthetic pigments and leaf area, resulting in less carbon assimilation. Additionally, metabolic activities being diverted for defense also contribute to decreased yield [56-58]. In the current study, mung bean cultivars exhibited reduced yield under salt stress, and the yield further reduced with higher levels of salt treatment. Similar findings of significant decline in yield under salinity stress were observed in various crops, such as wheat [59-64]. The presented study confirms that a higher degree of salt stress caused the maximum reduction in yield.
The increasing agricultural productivity improves world economy of the countries. In the present study, salt concentration caused a negative impact on the growth and yield of mung bean plants. Nigam also reported that the salt stress reduced the growth and yield of agricultural plants [31]. Despite that the application of exogenous protectants improves the plant growth and yield under various abiotic stress [41-42,31,65]. The higherprotection was seen in 50ppm salt +40%HA than 100ppm salt +40%HA. This means that HA will have a useful tool for agricultural sustainability against salt stress. PCAs analysis showed the application of HA and cultivars tolerance against salt stress based on strong relation with yield and plant physiological characteristics. PCA analysis of mung bean cultivar Pusa Baisakhi variate from PC1 (83.36%) and PC2 (13.05%) with Eigenvalue PC1 (13.33) and PC2 (2.08). Correlation between parameters of cultivar showed a strong relation with biomass accumulation and yield characters. Treatment wise 100ppm salt caused more severe effect than 50ppm salt. While HA application positively correlate with all selected parameters. The higher values of loading were seen at vegetative stages than juvenile stages its means plant cope up of growth at vegetative stage. In case of cultivar ML-131 following the similar trends of Pusa Baisakhi with age and treatments. While correlation of growth and productivity of Pusa Baisakhi is more than cultivar ML-131. Therefore, study suggested that Pusa Baisakhi is the salt tolerance cultivar it will prefer for rotation in salt prone area and also HA applied as a protectant during juvenile stage of cultivar growing in salt prone area for sustainable agriculture [66].
Conclusion
Soil salinity is the burning problem worldwide causes negative effect on plants especially agricultural crops. Presented study was conducted to assess effect of salinity stress on two cultivars of mung bean and to evaluate the possible effects of exogenous protectants to overcome the salinity stress. For this, presented study selected two concentration of humic acid as a protectant viz. 20 and 40%, to assess the accuracy of protectants in minimizing variable salt stress on selected cultivars. The effects of exogenous protectants on the physiological, biochemical and yield traits of mung bean cultivars at two NaCl concentrations (50 ppm and 100 ppm) were investigated. Obtain results shows that higher salinity (100 ppm) caused higher effect on both cultivars and application of humic acid play protective role against salt stress compared to control plants. Study also indicates that under 100ppm salt stress, higher yield reduction was seen in cultivar ML-131 (-26.35%) than cultivar Pusa Baisakhi (-23.65%), while increment of yield due to applied protectants was higher in cultivar ML-131 (65.95%) than cultivar Pusa Baisakhi (8.54%) in treatment T4 with 40%HAand 50 ppm salt.Therefore, study concluded that based on biomass and yield reduction Pusa Baisakhi is more tolerance to salt than ML- 131. Therefore, the study promoted that under salt stress conditions humic acid uses as a protectant and cultivar Pusa Baisakhi as a tolerance species will be useful tool for sustainable agriculture.
Acknowledgments
The authors wish to thank the Department of Environmental Science, Savitribai Phule Pune University, Pune, Maharashtra for the platform that enabled this interesting work.
Data Availability
All related data given in this article.
References
- Diacono, M., & Montemurro, F. (2015). Effectiveness of organic wastes as fertilizers and amendments in salt-affected soils. Agriculture, 5(2), 221-230.
- Parihar, P., Singh, S., Singh, R., Singh, V. P., & Prasad, S.M. (2015). Effect of salinity stress on plants and its tolerance strategies: a review. Environmental science and pollution research, 22, 4056-4075.
- Kaya C, Ashraf M, Dikilitas M, Tuna AL. (2013). Alleviation of salt stress-inducedadverse effects on maize plants by exogenous application of indoleacetic acid (IAA)and inorganic nutrients – A field trial. Aust J Crop Sci 7 (2), 249–254.
- Hussain, S., Zhang, J. H., Zhong, C., ZHU, L. F., CAO, X.C., YU, S. M., ... & JIN, Q. Y. (2017). Effects of salt stress on rice growth, development characteristics, and the regulating ways: A review. Journal of integrative agriculture, 16(11), 2357-2374.
- Siddiqui, M. N., Mostofa, M. G., Akter, M. M., Srivastava,K., Sayed, M. A., Hasan, M. S., & Tran, L. S. P. (2017).Impact of salt-induced toxicity on growth and yield-potential of local wheat cultivars: Oxidative stress and ion toxicity are among the major determinants of salt-tolerant capacity. Chemosphere, 187, 385-394.
- Wicke, B., Smeets, E., Dornburg, V., Vashev, B., Gaiser, T., Turkenburg, W., & Faaij, A. (2011). The global technical and economic potential of bioenergy from salt-affected soils. Energy & Environmental Science, 4(8), 2669-2681.
- Shrivastava, P., & Kumar, R. (2015). Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi journal of biological sciences, 22(2), 123-131.
- Munns, R., & Gilliham, M. (2015). Salinity tolerance of crops–what is the cost?. New phytologist, 208(3), 668-673.
- Mandal, S., Raju, R., Kumar, A., Kumar, P., & Sharma, P. C. (2018). Current status of research, technology response and policy needs of salt-affected soils in India–a review. J. Indian Soc. Coast. Agric. Res, 36(2).
- Negrão, S., Schmöckel, S. M., & Tester, M. J. A. O. B. (2017). Evaluating physiological responses of plants to salinity stress. Annals of botany, 119(1), 1-11.
- Tanveer, M., & Shabala, S. (2018). Targeting redox regulatory mechanisms for salinity stress tolerance in crops. Salinity Responses and Tolerance in Plants, Volume 1: Targeting Sensory, Transport and Signaling Mechanisms, 213-234.
- Chaudhary, I. J., & Rathore, D. (2020). Relative effectiveness of ethylenediurea, phenyl urea, ascorbic acid and urea in preventing groundnut (Arachis hypogaea L) crop from ground level ozone. Environmental Technology & Innovation, 19, 100963.
- Akram, N. A., Shafiq, F., & Ashraf, M. (2017). Ascorbic acid-a potential oxidant scavenger and its role in plant development and abiotic stress tolerance. Frontiers in plant science, 8, 238088.
- Shahbaz, M., Abid, A., Masood, A., & Waraich, E. A. (2017). Foliar-applied trehalose modulates growth, mineral nutrition, photosynthetic ability, and oxidative defense system of rice (Oryza sativa L.) under saline stress. Journal of Plant Nutrition, 40(4), 584-599.
- Melendrez, M. M. (2020). Humic acid: The science of humus and how it benefits soil. Eco Farming Daily.[Online] Available from: https://www. ecofarmingdaily. com/build-soil/ humus/humic-acid, 3, 265-277.
- Sangakkara, U. R., Hartwig, U. A., & Nösberger, J. (1996). Root and shoot development of Phaseolus vulgaris L.(French beans) as affected by soil moisture and fertilizer potassium. Journal of Agronomy and Crop Science, 177(3), 145-151.
- Maclachlan, S., & Zalik, S. (1963). Plastid structure, chlorophyll concentration, and free amino acid composition of a chlorophyll mutant of barley. Canadian Journal of Botany, 41(7), 1053-1062.
- DUXIWRY, A., & YENTSCII, C. (1956). Plankton pigment nomagraphs. J. Mar. Rcs, 15, 92-101.
- Flint, S. D., Jordan, P. W., & Caldwell, M. M. (1985). Plant protective response to enhanced UVâ?B radiation under field conditions: leaf optical properties and photosynthesis. Photochemistry and Photobiology, 41(1), 95-99.
- Keller, T., & Schwager, H. (1977). Air pollution and ascorbic acid. European Journal of Forest Pathology, 7(6), 338-350.
- Rahman, M. M., Rahman, M. A., Miah, M. G., Saha, S. R., Karim, M. A., & Mostofa, M. G. (2017). Mechanistic insight into salt tolerance of Acacia auriculiformis: the importance of ion selectivity, osmoprotection, tissue tolerance, and Na+ exclusion. Frontiers in plant science, 8, 240904.
- Akram, S., Siddiqui, M. N., Hussain, B. N., Al Bari, M. A.,Mostofa, M. G., Hossain, M. A., & Tran, L. S. P. (2017). Exogenous glutathione modulates salinity tolerance of soybean [Glycine max (L.) Merrill] at reproductive stage. Journal of Plant Growth Regulation, 36, 877-888.
- Acosta-Motos, J. R., Ortuño, M. F., Bernal-Vicente, A., Diaz-Vivancos, P., Sanchez-Blanco, M. J., & Hernandez, J. A. (2017). Plant responses to salt stress: adaptive mechanisms. Agronomy, 7(1), 18.
- Adolf, V. I., Shabala, S., Andersen, M. N., Razzaghi, F., & Jacobsen, S. E. (2012). Varietal differences of quinoa’s tolerance to saline conditions. Plant and Soil, 357, 117-129.
- Shahid, M. A., Sarkhosh, A., Khan, N., Balal, R. M., Ali, S., Rossi, L., ... & Garcia-Sanchez, F. (2020). Insights into the physiological and biochemical impacts of salt stress on plant growth and development. Agronomy, 10(7), 938.
- Elâ?Shafey, N. M., Avramova, V., Beemster, G. T., Korany,S. M., & AbdElgawad, H. (2023). B2O3 nanoparticles alleviate salt stress in maize leaf growth zones by enhancing photosynthesis and maintaining mineral and redox status. Physiologia Plantarum, 175(5), e14033.
- Souza, A. C., Olivares, F. L., Peres, L. E. P., Piccolo, A., & Canellas, L. P. (2022). Plant hormone crosstalk mediated by humic acids. Chemical and Biological Technologies in Agriculture, 9(1), 29.
- Daur, I., & Bakhashwain, A. A. (2013). Effect of humic acid on growth and quality of maize fodder production. Pak. J. Bot, 45(S1), 21-25.
- Muhammad, I., Shalmani, A., Ali, M., Yang, Q. H., Ahmad, H., & Li, F. B. (2021). Mechanisms regulating the dynamics of photosynthesis under abiotic stresses. Frontiers in plant science, 11, 615942.
- Kumar, S., Li, G., Yang, J., Huang, X., Ji, Q., Liu, Z., ... & Hou, H. (2021). Effect of salt stress on growth, physiological parameters, and ionic concentration of water dropwort (Oenanthe javanica) cultivars. Frontiers in plant science, 12, 660409.
- Nigam, B., Dubey, R. S., & Rathore, D. (2022). Protective role of exogenously supplied salicylic acid and PGPB (Stenotrophomonas sp.) on spinach and soybean cultivars grown under salt stress. Scientia Horticulturae, 293, 110654.
- Alam MA, Juraimi AS, Rafii MY, Hamid A (2015). Effect of salinity on biomass yield and physiological and stem-root anatomical characteristics of purslane (Portulaca oleracea L.) accessions. Biomed Res Int.
- Epub 2015 Feb 23. Retraction in: Biomed Res Int. 2019 Aug 8;2019:9458681. PMID: 25802833; PMCID: PMC4352753.
- Negrão, S., Schmöckel, S. M., & Tester, M. J. A. O. B. (2017). Evaluating physiological responses of plants to salinity stress. Annals of botany, 119(1), 1-11.
- Ali, Q., Shabaan, M., Ashraf, S., Kamran, M., Zulfiqar, U., Ahmad, M., ... & Arslan, M. (2023). Comparative efficacy of different salt tolerant rhizobial inoculants in improving growth and productivity of Vigna radiata L. under salt stress. Scientific Reports, 13(1), 17442.
- Song, X., Chen, M., Chen, W., Jiang, H., & Yue, X. (2020). Foliar application of humic acid decreased hazard of cadmium toxicity on the growth of Hybrid Pennisetum. Acta Physiologiae Plantarum, 42, 1-11.
- Cedeño-García, G. A., Gerding, M., Moraga, G., Inostroza, L., Fischer, S., Sepúlveda-Caamaño, M., & Oyarzúa, P. (2018). Plant growth promoting rhizobacteria with ACC deaminase activity isolated from Mediterranean dryland areas in Chile: Effects on early nodulation in alfalfa. Chilean journal of agricultural research, 78(3), 360-369.
- Souana, K., Taïbi, K., Abderrahim, L. A., Amirat, M., Achir, M., Boussaid, M., & Mulet, J. M. (2020). Salt-tolerance in Vicia faba L. is mitigated by the capacity of salicylic acid to improve photosynthesis and antioxidant response. Scientia Horticulturae, 273, 109641.
- Pour-Aboughadareh, A., Mehrvar, M. R., Sanjani, S., Amini, A., Nikkhah-Chamanabad, H., & Asadi, A. (2021). Effects of salinity stress on seedling biomass, physiochemical properties, and grain yield in different breeding wheat genotypes. Acta Physiologiae Plantarum, 43(7), 98.
- Zhao, S., Zhang, Q., Liu, M., Zhou, H., Ma, C., & Wang,P. (2021). Regulation of plant responses to salt stress.International Journal of Molecular Sciences, 22(9), 4609.
- Rathore, D., & Chaudhary, I. J. (2019). Ozone risk assessment of castor (Ricinus communis L.) cultivars using open top chamber and ethylenediurea (EDU). Environmental Pollution, 244, 257-269.
- Chaudhary, I. J., & Rathore, D. (2018). Suspended particulate matter deposition and its impact on urban trees. Atmospheric Pollution Research, 9(6), 1072-1082.
- Chaudhary, I. J., & Rathore, D. (2019). Dust pollution: Its removal and effect on foliage physiology of urban trees. Sustainable Cities and Society, 51, 101696.
- Chaudhary, I. J., & Rathore, D. (2021). Assessment of ozone toxicity on cotton (Gossypium hirsutum L.) cultivars: Its defensive system and intraspecific sensitivity. Plant Physiology and Biochemistry, 166, 912-927.
- Xu, Z., Jiang, Y., & Zhou, G. (2015). Response and adaptation of photosynthesis, respiration, and antioxidant systems to elevated CO2 with environmental stress in plants. Frontiers in plant science, 6, 701.
- Li, Y., He, N., Hou, J., Xu, L., Liu, C., Zhang, J., ... & Wu, X.(2018). Factors influencing leaf chlorophyll content in natural forests at the biome scale. Frontiers in Ecology and Evolution, 6, 64.
- Pareek, S., Sagar, N. A., Sharma, S., Kumar, V., Agarwal, T., Gonzálezâ?Aguilar, G. A., & Yahia, E. M. (2017). Chlorophylls: Chemistry and biological functions. Fruit and Vegetable Phytochemicals: Chemistry and Human Health, 2nd Edition, 269-284.
- HniliÄková, H., HniliÄka, F., Martinkova, J., & Kraus, K. (2017). Effects of salt stress on water status, photosynthesis and chlorophyll fluorescence of rocket.
- Kamran, M., Parveen, A., Ahmar, S., Malik, Z., Hussain, S., Chattha, M. S., ... & Chen, J. T. (2019). An overview of hazardous impacts of soil salinity in crops, tolerance mechanisms, and amelioration through selenium supplementation. International journal of molecular sciences, 21(1), 148.
- Müller, M., & Munné-Bosch, S. (2021). Hormonal impact on photosynthesis and photoprotection in plants. Plant Physiology, 185(4), 1500-1522.
- El-Sarkassy, N. M., Ibrahim, S. A., & Desoky, E. M. (2017). Salinity stress amelioration using humic acid and mycorrhizae on pepper plants. Zagazig Journal of Agricultural Research, 44(6), 2515-2527.
- Ennab, H. A., Mohamed, A. H., El-Hoseiny, H. M., Omar,A., Hassan, I. F., Gaballah, M. S., ... & Alam-Eldein, S.M. (2023). Humic acid improves the resilience to salinity stress of drip-irrigated Mexican lime trees in saline clay soils. Agronomy, 13(7), 1680.
- Sofi,A., Ebrahimi, M., & Shirmohammadi, E. (2021). Influence of humic acid on germination, morphological characteristics and photosynthesis pigments of Trifolium alexandrium L. under salinity stress. Ecopersia, 9(4), 287-297.
- Swapnil, P., Meena, M., Singh, S. K., Dhuldhaj, U. P., & Marwal, A. (2021). Vital roles of carotenoids in plants and humans to deteriorate stress with its structure, biosynthesis, metabolic engineering and functional aspects. Current Plant Biology, 26, 100203.
- El-Ghamry, A. M., Abd El-Hai, K. M., & Ghoneem, K. M. (2009). Amino and humic acids promote growth, yield and disease resistance of faba bean cultivated in clayey soil. Aust.J. Basic Appl. Sci, 3(2), 731-739.
- Caverzan A, Passaia G, Rosa SB, Ribeiro CW, Lazzarotto F and Margis-Pinheiro M (2012). Plant responses to stresses: roleof ascorbate peroxidase in the antioxidant protection. Genet Mol Biol 35:1011-1019.
- Hasanuzzaman, M., Alam, M. M., Nahar, K., Ahamed, K. U., & Fujita, M. (2014). Exogenous salicylic acid alleviates salt stress-induced oxidative damage in Brassica napus by enhancing the antioxidant defense and glyoxalase systems. Australian Journal of Crop Science, 8(4), 631-639.
- Chaudhary IJ, Nigam B, Rathore D. (2023). Effect of elevated ozone on soybean (Glycine max L.) cultivar: Role of orange juice and synthetic ascorbic acid. Nature Environment and Pollution Technology 22(3) 1225-1238.
- Batista, V. C. V., Pereira, I. M. C., de Oliveira Paula-Marinho,S., Canuto, K. M., Pereira, R. D. C. A., Rodrigues, T. H. S.,... & de Carvalho, H. H. (2019). Salicylic acid modulates primary and volatile metabolites to alleviate salt stress-induced photosynthesis impairment on medicinal plant Egletes viscosa. Environmental and Experimental Botany, 167, 103870.
- Kausar, F., Shahbaz, M., & Ashraf, M. (2013). Protective role of foliar-applied nitric oxide in Triticum aestivum under saline stress. Turkish Journal of Botany, 37(6), 1155-1165.
- Latef, A. A. H. A., & Chaoxing, H. (2011). Effect of arbuscular mycorrhizal fungi on growth, mineral nutrition, antioxidant enzymes activity and fruit yield of tomato grown under salinity stress. Scientia Horticulturae, 127(3), 228-233.
- Mostofa, M. G., Fujita, M., & Tran, L. S. P. (2015). Nitric oxide mediates hydrogen peroxide-and salicylic acid-induced salt tolerance in rice (Oryza sativa L.) seedlings. Plant growth regulation, 77, 265-277.
- Abdel Latef, A. A. H., & Chaoxing, H. (2014). Does inoculation with Glomus mosseae improve salt tolerance in pepper plants?. Journal of Plant Growth Regulation, 33, 644-653.
- Aminifard, M. H., Aroiee, H., Azizi, M., Nemati, H., & Jaafar,H. Z. (2012). Effect of humic acid on antioxidant activities and fruit quality of hot pepper (Capsicum annuum L.). Journal of Herbs, Spices & Medicinal Plants, 18(4), 360-369.
- Aminifard, M. H., Aroiee, H., Nemati, H., Azizi, M., & Khayyat, M. (2012). Effect of nitrogen fertilizer on vegetative and reproductive growth of pepper plants under field conditions. Journal of plant nutrition, 35(2), 235-242.
- Rathore, D., & Chaudhary, I. J. (2019). Ozone risk assessment of castor (Ricinus communis L.) cultivars using open top chamber and ethylenediurea (EDU). Environmental Pollution, 244, 257-269.