
Journal of Robotics and Automation Research(JRAR)
ISSN: 2831-6789 | DOI: 10.33140/JRAR
Impact Factor: 1.06

Research Article - (2025) Volume 6, Issue 1
Dsungaripterids: What Are They?
Received Date: Jan 15, 2025 / Accepted Date: Feb 13, 2025 / Published Date: Feb 17, 2025
Copyright: ©Â©2025 Carlos Albuquerque. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Citation: Albuquerque, C. (2025). Dsungaripterids: What Are They?. J Robot Auto Res, 6(1), 01-06.
Abstract
TDsungaripterids are a clade of pterodactyloid pterosaurs from the Early Cretaceous. Highly unusual, their affinities have proven problematic as some studies favour them as derived members of Germanodactylidae while others recover them as azhdarchoids, either at the base of the clade or nested deeper alongside Thalassodromidae. Here, following examples of recent re-evaluations of phylogenetic signals, I examine each individual character understood as evidence towards either result of dsungaripteroid phylogenetic placement. The overall results seem to lean towards an interpretation as derived germanodactylids, but several similarities with juvenile tapejarids may provide a scenario for azhdarchoid dsungaripterids.
Introduction
Dsungaripterids are a clade of pterodactyloid pterosaurs that lasted roughly the span of the Lower Cretaceous, Berriasian to Albian [1,2]. They can be considered highly aberrant by pterosaur standards, being characterised by thick bone walls, short wings, edentulous jaw tips and bone-encased teeth that are some of the most specialised known among sauropsids [2]. While some forms (i.e. Noripterus) have less extreme forms of these conditions, there isn’t a clear bridge between dsungaripteroids and other pterosaur groups. Due to this, interpreting their phylogenetic placement within Pterosauria has been a complicated process.
Traditionally, dsungaripterids have been linked with the Late Jurassic taxa Germanodactylus, Altmuehlopterus and Tendagu- rupterus among other, undescribed remains amidst others; see for a more comprehensive list) [2,3]. These taxa, usually grouped as Germanodactylidae, are rather close to the ancestral condition for Pterodactyloidea and even share cranial features with non-ptero- dactyloid monofenestratans like Darwinopterus, but bear several features in common with dsungaripterids¨ including robust denti- tion, the beginnings of jaw-tip edentuly in the form of an odontoid process, immensely expanded opisthotic processes and a strongly recurved femur. Under this interpretation, these pterosaurs repre- sent intermediate forms between generic pterodactyloids and the highly derived dsungaripterids [4].
A series of recent studies have however consistently recovered germanodactylids as close to ctenochasmatoids while dsungaripterids are part of the azhdarchoid assemblage amidst others) [5-7]. Few of these studies explore why such a relationship is recovered, usually just stopping at character datasets, but more recently examinations of Dsungaripterus ’ palate demonstrate several features previously thought to be restricted to Azhdarchoidea, strengthening this phylogenetic placement [8]. Previously, Kellner attempted to lump dsungaripterids, azhdarchoids and pteranodontians on the basis of all possessing a notarium, but several other researchers have pointed out that virtually all pterosaurs with wingspans under 2 meters lack it, rendering it an essentially pointless character (amidst others) [2,9,10]. While seldomly remarked, flapling tapejarids do bear a few unique cranial features rather similar to those seen in dsungaripterids (figure 1), including a nasoantorbital fenestra with a sub-triangular shape (a feature not seen in other pterosaurs, and quite opposite the condition seen in adult azhdarchoids where the nasoantorbital expands vastly) and expanded opisthotic processes, rendering the skull subtriangular in shape much as in dsungaripterids.
Under our current understanding of pterodactyloid phylogeny, both options are mutually exclusive (germanodactylids sit close to the pterodactyloid base, sometimes aligned with ctenochasmatoids in Archaeopterodactyloidea, while azhdarchoids are nested within the highly derived ornithocheiroid assemblage), and either option would reveal rather different evolutionary charts for not only Dsungaripteridae but potentially Pterodactyloidea as a whole. If dsungaripterids are germanodactylids, then they are ”basal” pterodactyloids that evolved from generalistic Jurassic forms and progressed in terms of speciation until their extinction in the mid- Cretaceous. If they are azhdarchoids, however, then they are part of an explosive adaptive radiation of Ornithocheiroidea in the earliest Cretaceous, the Berriasian alone seeing pterosaur faunas dominated by archaeopterodactyloids and non-pterodactyloids instantly replaced by highly derived ornithocheiroid taxa such as pteranodontians, tapejarids and even possible azhdarchids
with dsungaripterids potentially being an aberrant evolutionary experiment of this group [1].
Solving this dilemma would therefore provide insight to the evolution of Pterosauria during the Cretaceous, as well as assessing which characters are useful when considering phylogenetic studies. Pterosaur phylogenetic studies are noted as being rather complicated due to their rather derived characteristics being hard to interpret and map out so any additional clarity could prove vital in understanding their relationships and evolutionary history. Materials and Methods. I will examine each individual character cited in these phylogenetic analyses and explore on whereas they reflect genuine phylogenetic signals. A final statistical analysis accompanies this process. Bone reconstructions by corvarts (https://corvarts.wordpress.com/). References cited in text [11,12].
Results

Figure 1: Cranial comparisions between Dsungaripterus weii, Germanodactylus cristatus and juvenileSinopterus (’Nemicolopterus’) spp. [8,13]. Note shared characteristics among the three taxa (enlarged opisthotic proccesses, edentulism on at least the jaw tips), characters shared between only the first two (robust teeth) and characters shared between the first and last (subtriangular nasoantorbital).
• Jaw tip edentulism: Dsungaripterids represent a unique condition among pterosaurs between toothed snouts and a toothless beak, with 15 or less tooth pairs packed towards the base of the jaws while the distal margin is edentulous and covered in keratin. Many pterodactyloid clades possess an odontoid process (a keratin covered jaw tip that essentially acts as an additional tooth), including both ctenochasmatoids (i.e.Pterodactylus ) as well as istiodactylids odontoid processes also occur in a very few nonpterodactyloids, most notablyRhamphorhynchus [14]. Thus, the dsungaripterid condition is simply an extreme version of a trait observed in many pterosaur lineages, with the relatively large and straight odontoid processes in germanodactylids being possibly the ancestral condition. Because azhdarchoids are toothless the reacquisition of teeth would be less likely, but it is also possible that the last common ancestor between dsungaripterids and other azhdarchoids was toothed and might either have bore an intermediate condition as seen in dsungaripterids or simply a large odontoid process as seen in germanodactylids, leading to complete toothlessness in taxa like tapejarids and azhdarchids among others.
• Robust teeth: The teeth of dsungaripterids are unique among pterosaurs in being stout and encased in bone, rendering normal reptilian tooth replacement most likely impossible. This condition is extremely specialised among pterosaurs but could have had an ancestor in germanodactylid teeth, which while not atypical are already fairly robust compared to the teeth of contemporary pterodactyloids. Nonetheless, speciations towards durophagy have occured multiple times among monofenestratans, including wukongopterids the¨ bizarre ctenochasmatoid Cycnorhamphus and the azhdarchoidAlanqa, with the former having teeth convergent with those of germanodactylids. Still, while convergent evolution cannot be ruled out, only germanodactylids among pterodactyloids seem to have developed robust teeth, with both Cycnorhamphus and Alanqa (the latter being particularly significant as an azhdarchoid) having acquired jaw flanges to process hard items [2,15-17]. This renders a continuity between the robust teeth of germanodactylids and the aberrant bone-encased ones of dsungaripterids fairly plausible.
• Subtriangular nasoantorbital: In both dsungaripterids and flapling tapejarids the nasoantorbital fenetra expands downwards, forming a sub-triangular shape. This results in an upper jaw profile not seen in other pterosaur groups, with adult tapejarids even abandoning this condition and expanding the nasoantorbital above the eye socket. An azhdarchoid interpretation of Dsungaripteridae would be strongly supported if the condition seen in young tapejarids proved to be atavistic, since it is a highly unusual character that doesn’t seem to have convergently evolved multiple times. Germanodactylids lack this condition, having a medially expanding, suboval nasoantorbital like most early monofenestratans.
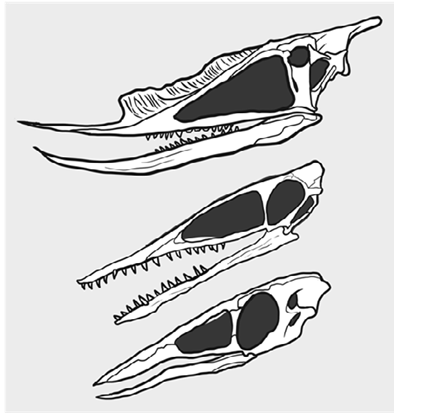
Figure 2: Palatal comparisons between Dsungaripterus weii (after), Caupedactylus ybaka andGermanodactylus cristatus [13]. Note similarities in the first two taxa in terms of complexity and fusion of pterygoid and ectopterygoid bones, while the latter has simpler, unfused elements more in line with the monofenestratan ancestral condition [8].
• Palate: The palate of dsungaripterids is most similar to that of azhdarchoids, sharing with them an ectopterygoid that overlays the pterygoid dorsally and merges with the pterygoid process posteriorly. This arrangement is not seen in other pterodactyloid clades (figure 2) and favours a relationship between dsungaripterids and azhdarchoids, particularly as the latter are seldomly durophagous like the former so convergent evolution would be less likely to explain this similarity. Further, the condition seen in dsungaripterids is closer to that seen in tapejarids than that seen in more derived azhdarchoids, suggesting that it represents a synapomorphy between both groups to the exclusion of other pterosaurs. Still, it does differ from the condition seen in tapejarids in that the ectopterygoid lies posterior to the edge of this palatal opening instead of reaching forward, so the possibility that it was acquired independently may not be ruled out yet [8].
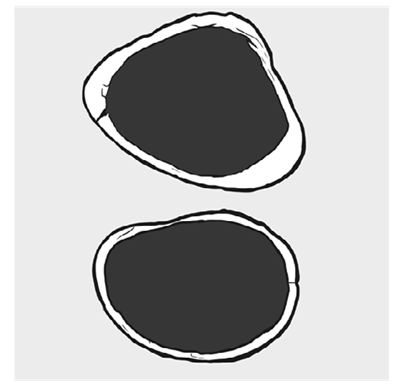
Figure 3: Bone wall comparisons between dsungaripterids and germanodactylids (SMF R 4915, after and azhdarchoids (Azhdarcho spp., after) [3,18]. In both dsungaripterids and germanodactylids the bone walls are exceptionally thick and show no evidence of pneumacy, while azhdarchoids lie at the polar opposite with millimetre thin bone walls with pneumatic foramen (not shown in cross-section).
• Bone wall thickness: Dsungaripterids have some of the thickest bone walls of any pterosaur, a trait they share with germanodactylids [3]. This stands in contrast to the rather thin bone walls of ornithocheiroids including azhdarchoids, often paper-thin [2]. Further, there appears to be no foramen allowing for bone pneumacy, rendering this condition exceptionally aberrant for an azhdarchoid but in line with a germanodactylid identity.
• Expanded opisthotic processes: Both germanodactylids (Witton among others) and juvenile tapejarids (figure 1) have expanded opisthotic processes that are less extreme than those seen in dsungaripterids, but are still substantially larger than those of other pterodactyloids. This likely reflects the presence of strong neck and jaw muscles and could easily have evolved multiple times. As adult tapejarids have less expanded opisthotic processes, it stands to reason that their presence in juveniles might be atavistic, potentially representing the condition between the last common ancestor of dsungaripterids and other azhdarchoids. In total, 2 characters (bone wall thickness and recurved femur) offer a strong phylogenetic signal for a germanodactylid dsungaripterids, 2 characters (nasoantorbital shape and palate morphology) offer a strong phylogenetic signal for azhdarchoid dsungaripterids, 1 character (robust teeth) offers a potential phylogenetic signal for germanodactylid dsungaripterids, 2 characters (expanded opisthotic processes and jaw tip endentulism) are of ambiguous
• Notarium: As noted previously, has attempted to argue that the notarium is a phylogenetic signal for Ornthocheiroidea, and posterior analyses citing dsungaripterids as azhdarchoids always use it as a character [9]. The problem is that the presence of the notarium is rather dependent on animal size: only pterosaurs with wingspans above 2 meters possess notaria, while all pterosaurs under this size value, including derived ornithocheiroids, lack it (Martin-Silverstone among others) [10]. Therefore, this character simply cannot provide a genuine phylogenetic signal, as it appears to be triggered by development. Its absence in some large pterosaurs (i.e. large ctenochasmatoids and rhamphorhynchids) may be of interest, but dsungaripteroids are notable for being rather heavy and relying on burst flights, so the presence of biomechanical reinforcements is a more plausible explanation rather than it reflecting a synapomorphy [2,3].
• Recurved femur: Both dsungaripterids and germanodactylids possess a strongly recurved femur. This feature is exclusive to them and not seen in any other pterosaur group, suggesting a genuine phylogenetic signal between both clades [2,3]. significance and 1 character (notarium) is effectively useless. A Germanodactylidae + Dsungaripteroidea clade is supported by 80% by the total quantitive analysis, while an azhdarchoid Dsungaripteroidea is supported by 65%; the final statistical analyses renders a support for the former in 70% and a support for the latter in 30%.
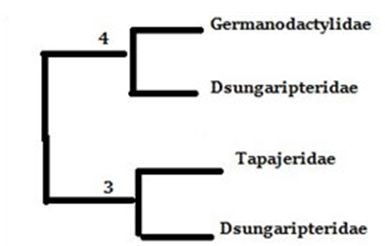
Figure 4: Phylogenetic tree constructed from the characters. As seen above, there is a slight favor towards a Dsungaripteridae + Germanodactylidae clade over an azhdarchoid identity for dsungaripterids.
Discussion
Dsungaripterids are thought to have occupied a highly specialised niche with no clear-cut modern analogue. They are largely terrestrial animals found in inland sites and seemed to have relied on burst flying with repeated landings, implicating them as ”poor flyers” [3]. Unlike modern fowl, however, they are traditionally interpreted as having been molluscivores, their edentulous jaw ends supposedly adapted for probing and their teeth suited to crack shells open. Still, unlike pterosaurs with known probing ecologies dsungaripterids appear to lack sensory pits and in fact their beaks are covered in hard keratin so this view could potentially be erroneous. Being terrestrial animals, it is likely that they were generalistic, albeit indeed with a durophagous bent, perhaps specialising in seeds, snails, turtles, scavenged bones or potentially even hard vegetation like cycads; it is worth noticing that dsungaripterids have proportionally large torsos compared to other pterosaurs, so suggestions of herbivory are not unreasonable, particularly as other pterosaurs like tapejarids have been suggested to have had frugivorous habits [2,3].
This rather atypical ecology has lead to their highly unusual morphology, seemingly prompting them to assume their more recognisable bauplan rather rapidly in the Berriasian with no known ”missing links”. If these animals are descendents from germanodactylids, then there is continuity with their less specialized Jurassic cousins, albeit one with several missing steps like the acquisition of more extensive edentulism, an azhdarchoid- like palate and subtriangular nasoantorbitals; this is in line with a gradual evolutionary process, with absenses explained either through gaps in the fossil reccord or bursts of rapid evolution. If dsungaripterids are azhdarchoids, however, their unique morphology was acquired exceptional fast, creating an entirely new bauplan radically different that that of its close relatives in the span of a few million years.
The placement of dsungaripterids has large consequences on the evolutionary history of Cretaceous pterosaurs. An adaptive radiation of some sort is recognised to have happened in the earliest Cretaceous, which lead to the replacement of at least rhamphorhynchids with pteranodontians as the main piscivorous and raptorial guilds and with the expansion of azhdarchoids into terrestrial ecologies [1,2]. Ctenochasmatoids and anurognathids are two major exceptions to this ornithocheiroid-dominated paradigm, showing that while abrupt this radiation still allowed some older clades to persist into the mid-Cretaceous. If dsungaripterids are germanodactylids, or some other form of basal pterodactyloid, this further demonstrates that pterosaur faunas of the Lower Cretaceous were mosaics of older taxa and ornithocheiroids, the former able to persist and thrive in spite of their new competitors.
If dsungaripterids are azhdarchoids, however, then ornithocheiroid radiation was far more explosive than previously understood, not only displacing the older germanodactylids but also branching into highly aberrant forms, dominating terrestrial ecologies even in the most specialised of niches. As such a rapid displacement eschews the conventions of competitive exclusion, it could imply a minor extinction event at the Jurassic/Cretaceous boundary that decimated several pterosaur clades prompting ornithocheiroids (and perhaps early birds) to expand into the vacant niches. This in turn might have ramifications for our understanding of pterosaur extinction, since Late Cretaceous pterosaur faunas are diverse but dominated by large sized ornithocheiroids [6].
The results of this paper lean towards an interpretation of dsungaripterids as germanodactylids, ergo following the more conventional scenario of several pterosaur lineages making it through the Jurassic-Cretaceous boundary and retaining some dominance even as ornithocheiroids expanded ecologically. However, future results may find more support for the explosive radiation of ornithocheiroids [19-21].
Conclusion
Analyses of both the various cited characters and their statistics lead to a stronger support for the traditional Germanodactylidae
+ Dsungaripteridae clade over dsungaripteroids being part of the azhdarchoid assemblage. However, a few ambiguous traits linking dsungaripterids to tapejarids remain, so further discoveries might reinforce the position of dsungaripteroids in Azhdarchoidea. Either result has rather drastic consequences for the structure of pterosaur faunas in the Cretaceous, the former indicating a mosaic between ornithocheiroids and earlier pterosaur lineages while the latter implies a rapid adaptive radiation on the part of azhdarchoids and ornithocheiroids and their replacement of other groups in response to some sort of faunal turnover.
References
- Dyke, G. J., Benton, M. J., Posmosanu, E., & Naish, D. (2010). Early Cretaceous (Berriasian) birds and pterosaurs from the Cornet bauxite mine, Romania. Palaeontology, 54(1), 79-95.
- Witton, M. P. (2013). Pterosaurs: natural history, evolution, anatomy. Princeton University Press.
- Fastnacht, M. (2005). The first dsungaripterid pterosaur from the Kimmeridgian of Germany and the biomechanics of pterosaur long bones. Acta Palaeontologica Polonica, 50(2).
- Lü, J., Unwin, D. M., Jin, X., Liu, Y., & Ji, Q. (2010). Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull. Proceedings of the Royal Society B: Biological Sciences, 277(1680), 383-389.
- Andres, B., & Qiang, J. (2008). A new pterosaur from the Liaoning Province of China, the phylogeny of the Pterodactyloidea, and convergence in their cervical vertebrae. Palaeontology, 51(2), 453-469.
- Longrich, N. R., Martill, D. M., & Andres, B. (2018). Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary. PLoS biology, 16(3), e2001663.
- Kellner, A. W., Weinschütz, L. C., Holgado, B., Bantim, R. A., & Sayao, J. M. (2019). A new toothless pterosaur (Pterodactyloidea) from Southern Brazil with insights into the paleoecology of a Cretaceous desert. Anais da Academia Brasileira de Ciências, 91(suppl 2), e20190768.
- He Chen, Shunxing Jiang, Alexander W.A. Kellner, Xin Cheng, Xinjun Zhang, Rui Qiu, Yang Li, Xiaolin Wang, New anatomical information on Dsungaripterus weii Young, 1964 with focus on the palatal region, Published April 1, 2020 PubMed 32274262
- Kellner, A. W. (2003). Pterosaur phylogeny and comments on the evolutionary history of the group. Geological Society, London, Special Publications, 217(1), 105-137.
- Bennett, C. S. (2001). The osteology and functional morphology of the Late Cretaceous pterosaur Pteranodon. Schweizerbart.
- Baron, M. G. (2021). The origin of Pterosaurs. Earth-Science Reviews, 221, 103777.
- Fernandes, D. L., Nunes, I., & Costa, F. R. (2021). A taxonomic approach on diagnostic characters used to define new pterosaur taxa and an estimation of pterosaur diversity. Anais da Academia Brasileira de Ciências, 93, e20201568.
- Bennett, S. C. (2006). Juvenile specimens of the pterosaur Germanodactylus cristatus, with a review of the genus. Journal of Vertebrate Paleontology, 26(4), 872-878.
- Martill, D. M. (2014). A functional odontoid in the dentary of the Early Cretaceous pterosaur Istiodactylus latidens: implications for feeding. Cretaceous Research, 47, 56-65.
- Lü, J., Xu, L., Chang, H., & Zhang, X. (2011). A new darwinopterid pterosaur from the Middle Jurassic of western Liaoning, northeastern China and its ecological implications. Acta Geologica Sinicaâ?English Edition, 85(3), 507-514.
- Naish, D., Witton, M. P., & Martin-Silverstone, E. (2021). Powered flight in hatchling pterosaurs: evidence from wing form and bone strength. Scientific Reports, 11(1), 13130.
- Martill, D. M., & Ibrahim, N. (2015). An unusual modification of the jaws in cf. Alanqa, a mid-Cretaceous azhdarchid pterosaur from the Kem Kem beds of Morocco. Cretaceous Research, 53, 59-67.
- Averianov, A., Dyke, G., Danilov, I., & Skutschas, P. (2015). The paleoenvironments of azhdarchid pterosaurs localities in the Late Cretaceous of Kazakhstan. ZooKeys, (483), 59.
- Martin-Silverstone, E., Witton, M. P., Arbour, V. M., & Currie, P. J. (2016). A small azhdarchoid pterosaur from the latest Cretaceous, the age of flying giants. Royal Society Open Science, 3(8), 160333.
- Martill, D. M., Smith, R. E., Longrich, N., & Brown, J. (2021). Evidence for tactile foraging in pterosaurs: a sensitive tip to the beak of Lonchodraco giganteus (Pterosauria, Lonchodectidae) from the Upper Cretaceous of southern England. Cretaceous Research, 117, 104637.
- Smith, R. E., Martill, D. M., Kao, A., Zouhri, S., & Longrich, N. (2021). A long-billed, possible probe-feeding pterosaur (Pterodactyloidea:? Azhdarchoidea) from the mid-Cretaceous of Morocco, North Africa. Cretaceous Research, 118, 104643.
- Smith, R. E., Chinsamy, A., Unwin, D. M., Ibrahim, N., Zouhri, S., & Martill, D. M. (2022). Small, immature pterosaurs from the Cretaceous of Africa: implications for taphonomic bias and palaeocommunity structure in flying reptiles. Cretaceous Research, 130, 105061.